El objetivo de este método es investigar cómo la entrada de retina influye en el crecimiento y desarrollo del tectum óptico, el centro de procesamiento visual en el cerebro del pez cebra. Al extraer un ojo y luego comparar los dos lados del tectum óptico, se pueden observar y normalizar los cambios técnicos dentro de la misma muestra, lo que permite la comparación entre múltiples muestras. Los enfoques moleculares modernos combinados con esta técnica proporcionarán información sobre los mecanismos subyacentes al crecimiento y desarrollo del sistema visual, así como a la degeneración y regeneración axonal.
Los sistemas sensoriales (visual, auditivo y somatosensorial) recopilan información de los órganos externos y transmiten esa información al sistema nervioso central, generando “mapas” del mundo externo a través delmesencéfalo 1,2. La visión es la modalidad sensorial dominante para casi todos los vertebrados, incluidos muchos peces. La retina, el tejido neural en el ojo, recopila información con un circuito neuronal que consiste principalmente en fotorreceptores, células bipolares y células ganglionares de la retina (RGC), las neuronas de proyección de la retina. Los RGC tienen axones largos que encuentran su camino a través de la superficie interna de la retina hasta la cabeza del nervio óptico, donde fasciculan y viajan juntos a través del cerebro, terminando finalmente en el centro de procesamiento visual en el mesencéfalo dorsal. Esta estructura se denomina tectum óptico en peces y otros vertebrados no mamíferos y es homóloga al colículo superior en mamíferos3.
El tectum óptico es una estructura multicapa bilateralmente simétrica en el mesencéfalo dorsal. En el pez cebra y la mayoría de los otros peces, cada lóbulo del tectum óptico recibe información visual únicamente del ojo contralateral, de modo que el nervio óptico izquierdo termina en el lóbulo tectal derecho y el nervio óptico derecho termina en el lóbulo tecttal izquierdo4 (Figura 1). Al igual que su contraparte en mamíferos, el colículo superior, el tectum óptico integra la información visual con otras entradas sensoriales, incluyendo la audición y la somatosensación, controlando los cambios en la atención visual y los movimientos oculares como las sacadas 1,5,6. Sin embargo, a diferencia del colículo superior de los mamíferos, el tectum óptico genera continuamente nuevas neuronas y glía a partir de un nicho especializado de células madre cerca de los bordes medial y caudal de los lóbulos tectales llamado zona de proliferacióntectal 7. El mantenimiento de progenitores proliferativos en el tectum óptico y otras regiones del sistema nervioso central contribuye, en parte, a la notable capacidad regenerativa documentada en el pez cebra8.
Trabajos previos que examinaron los cerebros de peces ciegos o tuertos revelaron que el tamaño del tectum óptico es directamente proporcional a la cantidad de inervación retiniana que recibe 9,10,11. En los peces de cueva adultos, cuyos ojos degeneran en la embriogénesis temprana, el tectum óptico es notablemente más pequeño que el de los peces de superficie avistados estrechamente relacionados9. La degeneración del ojo de pez de cueva se puede bloquear reemplazando la lente endógena con una lente de un pez de superficie durante la embriogénesis. Cuando estos peces de cueva tuertos se crían hasta la edad adulta, el lóbulo tectal inervado contiene aproximadamente un 10% más de células que el lóbulo tectal no inervado9. Del mismo modo, en los killifish larvales que fueron incubados con tratamientos químicos para generar ojos de diferentes tamaños dentro del mismo individuo, el lado del tectum con más inervación era más grande y contenía más neuronas10. La evidencia de los experimentos de aplastamiento del nervio óptico en peces de colores adultos indica que la inervación promueve la proliferación, con la proliferación de células tectales disminuyendo cuando se interrumpió la inervación11.
Confirmando y extendiendo estos estudios clásicos, varios informes recientes proporcionan datos que sugieren que la proliferación en respuesta a la inervación está modulada, al menos en parte, por la vía BDNF-TrkB12,13. Quedan muchas preguntas abiertas sobre el crecimiento y desarrollo del tectum óptico, incluida la forma en que un sistema sensorial en desarrollo hace frente a las lesiones y la degeneración del axón, qué señales celulares y moleculares permiten que la entrada de la retina regule el crecimiento del tectum óptico, cuándo estos mecanismos se activan y si la proliferación y la diferenciación vinculadas a la inervación permiten que la retina y su tejido objetivo coordinen las tasas de crecimiento y garanticen un mapeo retinotópico preciso. Además, hay preguntas mucho más grandes sobre el desarrollo dependiente de la actividad que se pueden abordar interrogando el sistema visual del pez cebra con enfoques quirúrgicos como el que se describe a continuación.
Para investigar los mecanismos celulares y moleculares por los cuales la actividad neuronal, específicamente a partir de la entrada visual, altera la supervivencia y proliferación celular, el enfoque descrito compara directamente los lóbulos tectales inervados y denervados (Figura 1) dentro de larvas individuales de pez cebra. Este método permite la documentación de la degeneración del axón RGC en el tectum óptico y la confirmación de que el número de células mitóticas se correlaciona con la inervación.
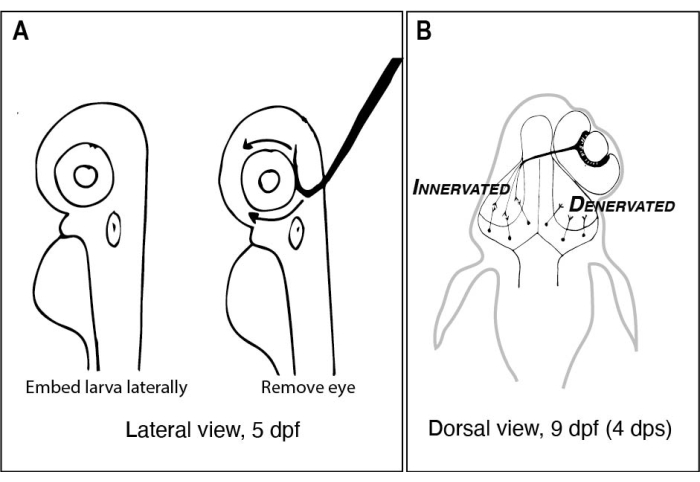
Figura 1: Bocetos de larvas de pez cebra antes y después de la extracción unilateral del ojo. (A) Dibujo de larvas de 5 dpf vistas bajo un microscopio de disección. Cada larva está incrustada en agarosa de bajo punto de fusión y orientada lateralmente antes de que se use una aguja de tungsteno con una punta afilada y enganchada para sacar el ojo hacia arriba (ojo izquierdo en este ejemplo). (B) Dibujo de la vista dorsal de una larva de 9 dpf resultante de la cirugía representada en A. Solo tres axones RGC altamente esquematizados del ojo derecho se muestran desfasciculando y conectando con neuronas en el lóbulo tectal izquierdo. Abreviaturas: dpf = días después de la fertilización; dps = días después de la cirugía; RGC = células ganglionares de la retina. Haga clic aquí para ver una versión más grande de esta figura.
