Målet med denna metod är att undersöka hur retinal inmatning påverkar tillväxten och utvecklingen av det optiska tectumet, det visuella bearbetningscentret i zebrafiskhjärnan. Genom att ta bort ett öga och sedan jämföra de två sidorna av det optiska tectumet kan tektala förändringar inom samma prov observeras och normaliseras, vilket möjliggör jämförelse över flera prover. Moderna molekylära tillvägagångssätt i kombination med denna teknik kommer att ge insikter i mekanismerna bakom visuell systemtillväxt och utveckling, liksom axonal degeneration och regenerering.
Sensoriska system – visuella, auditiva och somatosensoriska – samlar in information från yttre organ och vidarebefordrar den informationen till centrala nervsystemet och genererar “kartor” över den yttre världen över mitthjärnen 1,2. Vision är den dominerande sensoriska modaliteten för nästan alla ryggradsdjur, inklusive många fiskar. Näthinnan, den neurala vävnaden i ögat, samlar information med en neuronal krets som huvudsakligen består av fotoreceptorer, bipolära celler och retinala ganglionceller (RGC), näthinnans projektionsneuroner. RGC har långa axoner som hittar sin väg över näthinnans inre yta till synnervshuvudet, där de fascikulaterar och reser tillsammans genom hjärnan och slutligen slutar i det visuella bearbetningscentret i dorsala mitthjärnan. Denna struktur kallas optisk tectum hos fisk och andra icke-däggdjursryggradsdjur och är homolog mot den överlägsna colliculus hos däggdjur3.
Det optiska tectumet är en bilateralt symmetrisk flerskiktad struktur i dorsala mitthjärna. Hos zebrafiskar och de flesta andra fiskar får varje lob i det optiska tektumet visuell inmatning enbart från det kontralaterala ögat, så att vänster synnerv slutar i höger tektallob och höger synnerv slutar i vänster tektallob4 (Figur 1). Liksom sin däggdjurs motsvarighet, den överlägsna colliculus, integrerar det optiska tectum visuell information med andra sensoriska ingångar, inklusive audition och somatosensation, som kontrollerar förändringar i visuell uppmärksamhet och ögonrörelser som saccades 1,5,6. Men till skillnad från däggdjurets överlägsna colliculus genererar det optiska tectum kontinuerligt nya neuroner och glia från en specialiserad stamcellsnisch nära de mediala och kaudala kanterna på tektalloberna som kallas tectal proliferationszon7. Upprätthållande av proliferativa förfäder i det optiska tectumet och andra regioner i centrala nervsystemet bidrar delvis till den anmärkningsvärda regenerativa kapacitet som dokumenteras i zebrafisk8.
Tidigare arbete som undersökte hjärnorna hos blinda eller enögda fiskar avslöjade att optisk tektumstorlek är direkt proportionell mot mängden retinal innervation den får 9,10,11. Hos vuxna grottfiskar, vars ögon degenererar i tidig embryogenes, är det optiska tektumet märkbart mindre än för närstående, synad ytfisk9. Grottfiskögondegeneration kan blockeras genom att ersätta den endogena linsen med en lins från en ytfisk under embryogenesen. När dessa enögda grottfiskar föds upp till vuxen ålder innehåller den innerverade tektalloben cirka 10% fler celler än den icke-innerverade tektalloben9. På samma sätt, i larvdödfisk som inkuberades med kemiska behandlingar för att generera ögon av olika storlekar inom samma individ, var sidan av tectum med mer innervation större och innehöll fler neuroner10. Bevis från optiska nervkrossexperiment hos vuxna guldfiskar indikerar att innervation främjar proliferation, med tektal cellproliferation som minskar när innervation stördes11.
Bekräftar och utvidgar dessa klassiska studier, flera nya rapporter ger data som tyder på att proliferation som svar på innervation moduleras, åtminstone delvis, av BDNF-TrkB-vägen12,13. Många öppna frågor om optisk tektumtillväxt och utveckling kvarstår, inklusive hur ett utvecklande sensoriskt system hanterar skada och axondegenerering, vilka cellulära och molekylära signaler möjliggör retinal ingång för att reglera optisk tektumtillväxt, när dessa mekanismer blir aktiva och om innervationskopplad proliferation och differentiering gör det möjligt för näthinnan och dess målvävnad att samordna tillväxthastigheter och säkerställa korrekt retinotopisk kartläggning. Dessutom finns det mycket större frågor om aktivitetsberoende utveckling som kan hanteras genom att förhöra zebrafiskens visuella system med kirurgiska tillvägagångssätt som den som beskrivs nedan.
För att undersöka de cellulära och molekylära mekanismer genom vilka neural aktivitet, specifikt från visuell inmatning, förändrar cellöverlevnad och proliferation, jämför det beskrivna tillvägagångssättet direkt innerverade och denerverade tektallober (Figur 1) inom enskilda zebrafisklarver. Denna metod möjliggör dokumentation av RGC-axondegenerering i det optiska tektumet och bekräftelse på att antalet mitotiska celler korrelerar med innervation.
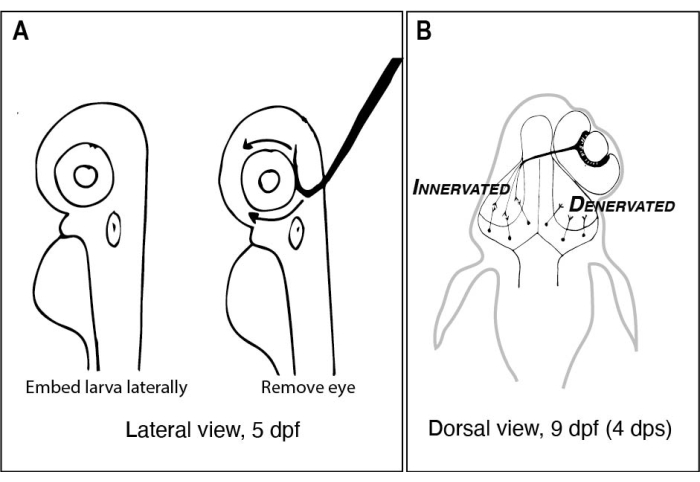
Figur 1: Skisser av zebrafisklarver före och efter ensidig ögonborttagning. (A) Ritning av 5 dpf larver sett under ett dissekerande mikroskop. Varje larv är inbäddad i lågsmältpunkt agaros och orienterad i sidled innan en volframnål med en skarp, krokad spets används för att skopa ut ögat uppåt (vänster öga i detta exempel). (B) Ritning av den dorsala vyn av en 9 dpf larv som härrör från den operation som avbildas i A. Endast tre mycket schematiserade RGC-axoner från höger öga visas defascikulerande och anslutande med neuroner i vänster tektallob. Förkortningar: dpf = dagar efter befruktning; dps = dagar efter operationen; RGC = retinala ganglionceller. Klicka här för att se en större version av denna figur.
