النيوترون هو جسيم ضخم بدون شحنة تم استخدامه بنجاح على مر السنين لفحص العينات في مختلف المجالات من الفيزياء الأساسية إلى علم الأحياء1. بالنسبة للتطبيقات البيولوجية ، يتم استخدام تشتت النيوترونات بزاوية صغيرة ، وتشتت النيوترونات غير المرن ، وعلم البلورات النيوتروني وقياس الانعكاس على نطاق واسع2،3،4. يوفر تشتت النيوترونات غير المرن قياسا متوسطا للديناميكيات دون الحاجة إلى وضع علامات محددة في حد ذاتها ، وجودة إشارة لا تعتمد على الحجم أو البروتين5. يمكن إجراء القياس باستخدام بيئة معقدة للغاية للبروتين قيد الدراسة الذي يحاكي الوسط داخل الخلايا ، مثل المحللة البكتيرية المنزوعة أو حتى في الجسم الحي3،6،7. يمكن استخدام إعدادات تجريبية مختلفة لدراسة الديناميات ، وهي i) وقت الرحلة الذي يمنح الوصول إلى ديناميكيات ps-ps الفرعية ، ii) الوصول إلى التشتت العكسي إلى ديناميكيات ps-ns ، و iii) الوصول إلى الصدى المغزلي إلى الديناميكيات من ns إلى مئات ns. يستخدم التشتت العكسي للنيوترونات قانون براغ 2d sinθ = nλ ، حيث d هي المسافة بين المستويات في البلورة ، و θ زاوية التشتت ، و n ترتيب التشتت ، و λ الطول الموجي. يسمح استخدام البلورات للتشتت العكسي نحو أجهزة الكشف بتحقيق دقة عالية في الطاقة ، عادة ~ 0.8 μeV. لقياس تبادل الطاقة ، يتم استخدام إما محرك دوبلر يحمل بلورة في التشتت العكسي لتحديد وضبط الطول الموجي النيوتروني الوارد8،9،10 (الشكل 1) ، أو يمكن استخدام إعداد وقت الرحلة على حساب انخفاض في دقة الطاقة11.
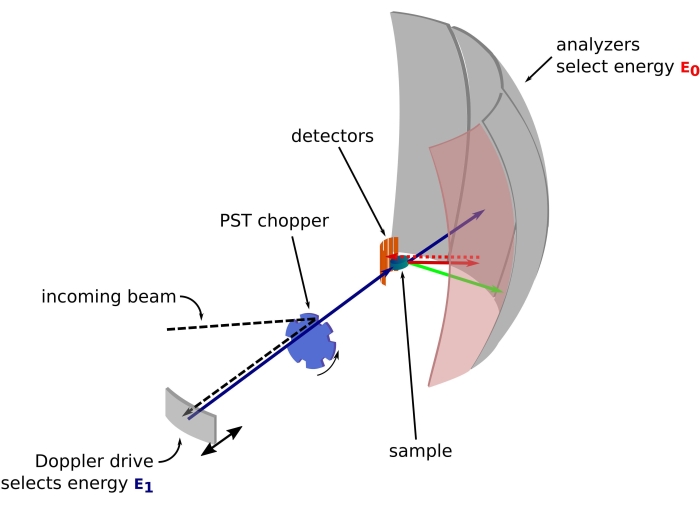
الشكل 1: رسم مطياف التشتت العكسي للنيوترونات بمحرك دوبلر. يصطدم الشعاع الوارد بمروحية تحويل فضاء الطور (PST)42 ، مما يزيد من التدفق في موضع العينة. ثم يتم تشتيتها مرة أخرى نحو العينة بواسطة محرك دوبلر ، الذي يختار الطاقة E1 (السهم السماوي). ثم يتم تشتيت النيوترونات بواسطة العينة (مع طاقات مختلفة ممثلة بلون الأسهم) والمحللين ، المصنوع من بلورات Si 111 ، سوف ينتشر فقط النيوترونات ذات الطاقة المحددة E0 (الأسهم ذات اللون الأحمر هنا). ومن ثم ، يتم الحصول على نقل الزخم q من الموضع المكتشف للنيوترون على صفيف الكاشف ، ويتم الحصول على نقل الطاقة من الفرق E1– E0. يتم استخدام وقت الرحلة المتوقع لنبضة النيوترون التي تنتجها PST لتجاهل الإشارة من النيوترونات المنتشرة مباشرة نحو أنابيب الكاشف. اختصار: PST = تحويل مساحة الطور. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
بالنسبة للتحليل الطيفي للتشتت العكسي ، تأتي المساهمة الرئيسية في الإشارة من العينات الغنية ببروتون الهيدروجين ، مثل البروتينات ، من التشتت غير المترابط ، حيث تظهر شدة التشتت Sinc (q ، ω) بواسطة Eq (1) 12
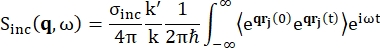 (1)
(1)
حيث σinc هو المقطع العرضي غير المترابط للعنصر المعتبر ، k ‘هو معيار المتجه الموجي المتناثر ، k هو معيار المتجه الموجي الوارد ، q (= k – k’) نقل الزخم ، r j (t) متجه موضع الذرة j في الوقت t ، و ω التردد المقابل لنقل الطاقة بين النيوترون الوارد والنظام. تشير الأقواس الزاوية إلى متوسط المجموعة. ومن ثم ، فإن التشتت غير المترابط يسبر الارتباط الذاتي للجسيم الواحد متوسط المجموعة لمواقع الذرة مع الوقت ويعطي متوسط الديناميكيات الذاتية على جميع الذرات في النظام وأصول زمنية مختلفة (متوسط المجموعة). دالة التشتت هي تحويل فورييه في زمن دالة التشتت الوسيطة I (q، t) ، والتي يمكن اعتبارها تحويل فورييه في الفضاء لدالة ارتباط فان هوف الموضحة بواسطة Eq (2):
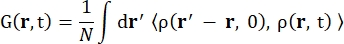 (2)
(2)
حيث ρ (r، t) هي كثافة احتمال إيجاد ذرة في الموضع r والوقت t 13.
بالنسبة لعملية الانتشار الويكي ، تنتج دالة الانتشار الذاتي (انظر Eq (3)) بعد تحويل فورييه المزدوج في دالة تشتت تتكون من لورنتزيان بعرض الخط المعطى بواسطة γ = Dq2.
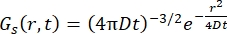 (3)
(3)
تم تطوير نماذج أكثر تطورا ووجدت مفيدة مثل نموذج انتشار القفز بواسطة Singwi و Sjölander لديناميات البروتين الداخليةps-ns 14 أو نموذج الدوران بواسطة Sears لمياه الترطيب15،16،17.
على أداة التشتت العكسي للنيوترونات (NBS) IN16B 8,9 في ILL، غرونوبل، فرنسا (الشكل التكميلي S1)، يتكون الإعداد الذي يشيع استخدامه مع البروتينات من بلورات Si 111 للمحللات مع محرك دوبلر لضبط الطول الموجي الوارد (الشكل التكميلي S2A)، مما يتيح الوصول إلى نطاق نقل الزخم ~ 0.2 Å-1 < q < ~ 2 Å-1 ونطاق نقل الطاقة -30 μeV < 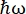 < 30 μeV المقابلة لجداول زمنية تتراوح من بضعة ps إلى عدد قليل من ns ومسافات قليلة Å. بالإضافة إلى ذلك ، يوفر IN16B إمكانية إجراء عمليات مسح مرنة وغير مرنة للنافذة الثابتة (E / IFWS) 10 ، والتي تشمل الحصول على البيانات عند نقل الطاقة الثابتة. نظرا لأن التدفق محدود عند العمل مع النيوترونات ، فإن E / IFWS يسمح بتعظيم التدفق لنقل طاقة واحد ، وبالتالي تقليل وقت الاكتساب اللازم للحصول على نسبة إشارة إلى ضوضاء مرضية. والخيار الأحدث هو النمط11 لمطياف التشتت العكسي ووقت الطيران (BATS) ، والذي يسمح بقياس مجموعة واسعة من عمليات نقل الطاقة ، (على سبيل المثال ، -150 μeV < < 150 μeV) ، مع تدفق أعلى من محرك دوبلر ، ولكن على حساب دقة طاقة أقل (الشكل التكميلي S2B).
< 30 μeV المقابلة لجداول زمنية تتراوح من بضعة ps إلى عدد قليل من ns ومسافات قليلة Å. بالإضافة إلى ذلك ، يوفر IN16B إمكانية إجراء عمليات مسح مرنة وغير مرنة للنافذة الثابتة (E / IFWS) 10 ، والتي تشمل الحصول على البيانات عند نقل الطاقة الثابتة. نظرا لأن التدفق محدود عند العمل مع النيوترونات ، فإن E / IFWS يسمح بتعظيم التدفق لنقل طاقة واحد ، وبالتالي تقليل وقت الاكتساب اللازم للحصول على نسبة إشارة إلى ضوضاء مرضية. والخيار الأحدث هو النمط11 لمطياف التشتت العكسي ووقت الطيران (BATS) ، والذي يسمح بقياس مجموعة واسعة من عمليات نقل الطاقة ، (على سبيل المثال ، -150 μeV < < 150 μeV) ، مع تدفق أعلى من محرك دوبلر ، ولكن على حساب دقة طاقة أقل (الشكل التكميلي S2B).
من الخصائص المهمة لتشتت النيوترونات أن المقطع العرضي غير المترابط σinc له قيمة أعلى 40 مرة للهيدروجين من الديوتيريوم ولا يكاد يذكر بالنسبة للعناصر الأخرى التي توجد عادة في العينات البيولوجية. لذلك ، يمكن دراسة ديناميكيات البروتينات في بيئة سائلة باستخدام مخزن مؤقت مقسم ، وتسمح حالة المسحوق بدراسة إما الديناميات الداخلية للبروتين مع مسحوق البروتين المهدرج المبلل ب D 2 O ، أودراسة ماء الترطيب لمسحوق البروتين perdeuterated المائي باستخدام H2O. في الحالة السائلة ، يسمح التشتت العكسي للنيوترونات عادة بالوصول في وقت واحد إلى الانتشار الذاتي لمركز الكتلة للبروتينات (الانتشار من نوع Fickian) وديناميكياتها الداخلية. هذه الأخيرة هي حركات العمود الفقري والسلسلة الجانبية التي يصفها عادة ما يسمى نموذج انتشار القفز أو غيره 3,18. في مساحيق البروتين المهدرجة ، يكون انتشار البروتين غائبا ويجب نمذجة الديناميات الداخلية فقط. بالنسبة لمياه الإماهة ، فإن مساهمات الحركات الانتقالية والدورانية لجزيئات الماء تقدم اعتمادا مختلفا على نقل الزخم q ، مما يسمح بتمييزها في عملية تحليل البيانات17.
توضح هذه الورقة طريقة التشتت العكسي للنيوترونات مع دراسة البروتينات التي وجد أنها قادرة على التكشف ، وتتجمع في شكل قانوني يتكون من أكوام من خيوط β – ما يسمى بنمط β المتقاطع19،20 – وتشكل أليافا ممدودة. هذا هو ما يسمى بتجميع الأميلويد ، والذي تمت دراسته على نطاق واسع بسبب دوره المركزي في الاضطرابات التنكسية العصبية مثل مرض الزهايمر أو مرض باركنسون21,22. إن دراسة بروتينات الأميلويد مدفوعة أيضا بالدور الوظيفي الذي يمكن أن تلعبه 23,24 أو إمكاناتها العالية لتطوير مواد حيوية جديدة25. لا تزال المحددات الفيزيائية والكيميائية لتجميع الأميلويد غير واضحة ، ولا تتوفر نظرية عامة لتجميع الأميلويد ، على الرغم من التقدم الهائل خلال السنوات الماضية21,26.
ينطوي تجميع الأميلويد على تغييرات في بنية البروتين واستقراره مع مرور الوقت ، والتي تنطوي دراستها بشكل طبيعي على ديناميكيات مرتبطة باستقرار تكوين البروتين ، ووظيفة البروتين ، ومشهد طاقة البروتين27. ترتبط الديناميكيات ارتباطا مباشرا باستقرار حالة معينة من خلال المساهمة الإنتروبية لأسرع الحركات28 ، ويمكن الحفاظ على وظيفة البروتين من خلال الحركات على نطاقات زمنية مختلفة من sub-ps للبروتينات الحساسة للضوء29 إلى ms لحركات المجال ، والتي يمكن تسهيلها بواسطة ديناميكيات بيكو ثانيةنانو ثانية 30.
سيتم تقديم مثالين على استخدام التحليل الطيفي للتشتت العكسي للنيوترونات لدراسة بروتينات الأميلويد ، أحدهما في الحالة السائلة لدراسة ديناميكيات البروتين والآخر في حالة المسحوق المائي لدراسة ديناميكيات الماء المماهة. يتعلق المثال الأول بتجميع الليزوزيم في كرات بحجم ميكرومتر (تسمى الجسيمات) متبوعة في الوقت الفعلي5 ، والثاني مقارنة لديناميكيات المياه في الحالات الأصلية والمجمعة للبروتين البشري تاو31.
الليزوزيم هو إنزيم يشارك في الدفاع المناعي ويتكون من 129 من بقايا الأحماض الأمينية. يمكن أن يشكل الليزوزيم جسيمات في المخزن المؤقت المثبط عند pD 10.5 وعند درجة حرارة 90 درجة مئوية. مع تشتت النيوترونات ، أظهرنا أن التطور الزمني لمعامل انتشار مركز الكتلة للليزوزيم يتبع الحركية الأسية المفردة لتألق ثيوفلافين T (مسبار فلوري يستخدم لمراقبة تكوين أنماط β المتقاطع الأميلويد32) ، مما يشير إلى أن الهياكل الفوقية للجسيمات وأنماط β المتقاطع تحدث في خطوة واحدة بنفس المعدل. علاوة على ذلك ، ظلت الديناميات الداخلية ثابتة طوال عملية التجميع ، والتي يمكن تفسيرها إما من خلال تغيير توافقي سريع لا يمكن ملاحظته على أدوات NBS ، أو من خلال عدم وجود تغيير كبير في الطاقة الداخلية للبروتين عند التجميع.
البروتين البشري تاو هو بروتين مضطرب جوهريا (IDP) يتكون من 441 من الأحماض الأمينية لما يسمى ب 2N4R isoform ، والذي يشارك بشكل خاص في مرض الزهايمر33. باستخدام التشتت العكسي للنيوترونات على مساحيق بروتين تاو perdeuterated ، أظهرنا أن ديناميكيات الماء المائيات تزداد في حالة الألياف ، مع وجود عدد أكبر من جزيئات الماء التي تخضع لحركات متعدية. تشير النتيجة إلى أن الزيادة في إنتروبيا الماء المائي قد تؤدي إلى رجفان الأميلويد في تاو.
