Het neutron is een ladingloos en massief deeltje dat in de loop der jaren met succes is gebruikt om monsters te onderzoeken op verschillende gebieden, van fundamentele fysica tot biologie1. Voor biologische toepassingen worden neutronenverstrooiing met een kleine hoek, inelastische neutronenverstrooiing en neutronenkristallografie en reflectometrie op grote schaal gebruikt 2,3,4. Inelastische neutronenverstrooiing biedt een ensemblegemiddelde meting van de dynamiek zonder dat specifieke etikettering per se nodig is, en een signaalkwaliteit die niet afhankelijk is van de grootte of het eiwit5. De meting kan worden uitgevoerd met behulp van een zeer complexe omgeving voor het onderzochte eiwit dat het intracellulaire medium nabootst, zoals een gedeutereerd bacterieel lysaat of zelfs in vivo 3,6,7. Verschillende experimentele opstellingen kunnen worden gebruikt om de dynamica te bestuderen, namelijk i) time-of-flight-giving access to sub-ps-ps dynamics, ii) backscattering-giving access to ps-ns dynamics, and iii) spin-echo-giving access to dynamics from ns to hundreds of ns. Neutronenverstrooiing maakt gebruik van de wet van Bragg 2d sinθ = nλ, waarbij d de afstand is tussen vlakken in een kristal, θ de verstrooiingshoek, n de verstrooiingsvolgorde en λ de golflengte. Het gebruik van kristallen voor backscattering naar de detectoren zorgt voor het bereiken van een hoge resolutie in energie, meestal ~ 0,8 μeV. Om de energie-uitwisseling te meten, wordt ofwel een Doppler-aandrijving met een kristal in backscattering gebruikt om de inkomende neutronengolflengte 8,9,10 te definiëren en af te stemmen (figuur 1), of een time-of-flight-opstelling kan worden gebruikt ten koste van een afname van de energieresolutie 11.
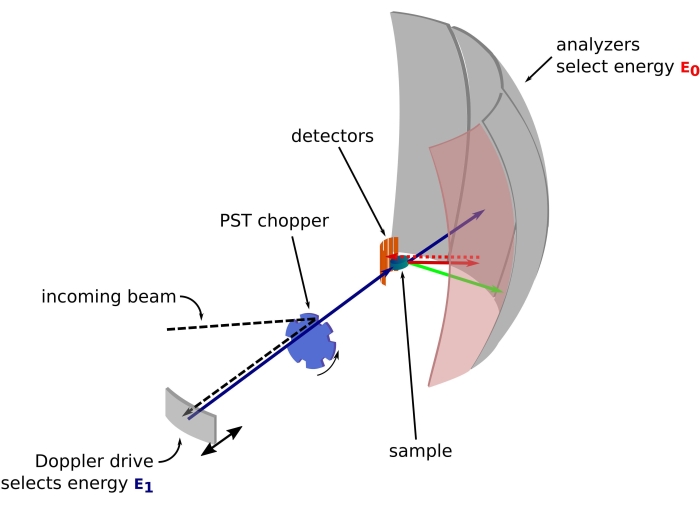
Figuur 1: Schets van een neutronen-backscattering spectrometer met een Doppler-aandrijving. De inkomende bundel raakt de phase space transformation (PST) chopper42, waardoor de flux op de monsterpositie toeneemt. Vervolgens wordt het teruggekaatst naar het monster door de Doppler-aandrijving, die een energie E1 (cyaanpijl) selecteert. De neutronen worden vervolgens verstrooid door het monster (met verschillende energieën vertegenwoordigd door de kleur van de pijlen) en de analyzers, gemaakt van Si 111-kristallen, zullen alleen neutronen met een specifieke energie E0 terugverstrooien (rood gekleurde pijlen hier). Daarom wordt de momentumoverdracht q verkregen uit de gedetecteerde positie van het neutron op de detectorarray en wordt de energieoverdracht verkregen uit het verschil E1– E0. De verwachte vluchttijd voor de neutronenpuls die door de PST wordt geproduceerd, wordt gebruikt om het signaal van de neutronen die rechtstreeks naar de detectorbuizen zijn verspreid, weg te gooien. Afkorting: PST = phase space transformation. Klik hier om een grotere versie van deze figuur te bekijken.
Voor backscatteringspectroscopie is de belangrijkste bijdrage aan het signaal van waterstofprotonrijke monsters, zoals eiwitten, afkomstig van onsamenhangende verstrooiing, waarvoor de verstrooiingsintensiteit Sinc(q, ω) wordt weergegeven door Eq (1)12
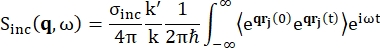 (1)
(1)
Waarbij σinc de onsamenhangende doorsnede van het beschouwde element is, is k’ de norm van de verstrooide golfvector, k de norm van de inkomende golfvector, q (= k – k’) de impulsoverdracht, r j(t) de positievector van atoom j op tijdstip t, en ω de frequentie die overeenkomt met de energieoverdracht tussen het inkomende neutron en het systeem. De hoekige haakjes geven het ensemblegemiddelde aan. Vandaar dat onsamenhangende verstrooiing de ensemble-gemiddelde zelfcorrelatie van één deeltje van atoomposities met de tijd onderzoekt en de zelfdynamiek gemiddeld geeft over alle atomen in het systeem en verschillende tijdsoorsprongen (ensemblegemiddelde). De verstrooiingsfunctie is de Fouriertransformatie in de tijd van de tussenliggende verstrooiingsfunctie I(q, t), die kan worden gezien als de Fouriertransformatie in de ruimte van de van Hove-correlatiefunctie weergegeven door Eq (2):
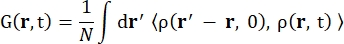 (2)
(2)
Waarbij ρ(r,t) de kansdichtheid is van het vinden van een atoom op positie r en tijd t 13.
Voor een Fickisch diffusieproces resulteert de zelfdiffusiefunctie (zie Eq (3)) na een dubbele Fouriertransformatie in een verstrooiingsfunctie bestaande uit een Lorentzian met een lijnbreedte gegeven door γ = Dq2.
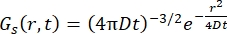 (3)
(3)
Meer geavanceerde modellen werden ontwikkeld en nuttig bevonden, zoals het sprongdiffusiemodel van Singwi en Sjölander voor ps-ns interne eiwitdynamica14 of het rotatiemodel van Sears voor hydratatiewater15,16,17.
Op het neutron backscattering (NBS) instrument IN16B 8,9 bij de IBL, Grenoble, Frankrijk (aanvullende figuur S1), bestaat een opstelling die vaak wordt gebruikt met eiwitten uit Si 111-kristallen voor de analyzers met een doppleraandrijving voor het afstemmen van de inkomende golflengte (aanvullende figuur S2A), waardoor toegang wordt gegeven tot het momentumoverdrachtsbereik ~ 0,2 Å-1 < q < ~ 2 Å-1 en energieoverdrachtsbereik van –30 μeV < 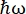 < 30 μeV-overeenkomend met tijdschalen variërend van enkele ps tot enkele ns en afstanden van enkele Å. Daarnaast biedt IN16B de mogelijkheid om elastische en inelastische fixed-window scans (E/IFWS)10 uit te voeren, waaronder data-acquisitie bij een vaste energieoverdracht. Omdat de flux beperkt is bij het werken met neutronen, maakt E/IFWS het mogelijk om de flux voor één energieoverdracht te maximaliseren, waardoor de acquisitietijd die nodig is om een bevredigende signaal-ruisverhouding te verkrijgen, wordt verkort. Een recentere optie is de backscattering en time-of-flight spectrometer (BATS) -modus11, waarmee een breed scala aan energieoverdrachten kan worden gemeten (bijv. -150 μeV < < 150 μeV), met een hogere flux dan met de Doppler-aandrijving, maar ten koste van een lagere energieresolutie (aanvullende figuur S2B).
< 30 μeV-overeenkomend met tijdschalen variërend van enkele ps tot enkele ns en afstanden van enkele Å. Daarnaast biedt IN16B de mogelijkheid om elastische en inelastische fixed-window scans (E/IFWS)10 uit te voeren, waaronder data-acquisitie bij een vaste energieoverdracht. Omdat de flux beperkt is bij het werken met neutronen, maakt E/IFWS het mogelijk om de flux voor één energieoverdracht te maximaliseren, waardoor de acquisitietijd die nodig is om een bevredigende signaal-ruisverhouding te verkrijgen, wordt verkort. Een recentere optie is de backscattering en time-of-flight spectrometer (BATS) -modus11, waarmee een breed scala aan energieoverdrachten kan worden gemeten (bijv. -150 μeV < < 150 μeV), met een hogere flux dan met de Doppler-aandrijving, maar ten koste van een lagere energieresolutie (aanvullende figuur S2B).
Een belangrijke eigenschap van neutronenverstrooiing is dat de onsamenhangende doorsnede σinc een 40 keer hogere waarde heeft voor waterstof dan voor deuterium en verwaarloosbaar is voor andere elementen die vaak in biologische monsters worden aangetroffen. Daarom kan de dynamiek van eiwitten in een vloeibare omgeving worden bestudeerd met behulp van een gedeutereerde buffer, en de poedertoestand maakt de studie mogelijk van ofwel eiwitinterne dynamiek met gehydrogeneerd eiwitpoeder gehydrateerd met D 2 O, of de studie van hydratatiewater voor geperdeutereerd eiwitpoeder gehydrateerdmet H2O. In vloeibare toestand maakt neutronenverstrooiing meestal gelijktijdige toegang mogelijk tot het massacentrum zelfdiffusie van eiwitten (Fickian-type diffusie) en hun interne dynamiek. De laatste zijn ruggengraat- en zijketenbewegingen die meestal worden beschreven door het zogenaamde sprongdiffusiemodel of andere 3,18. In gehydrogeneerde eiwitpoeders is de eiwitdiffusie afwezig en hoeft alleen de interne dynamiek te worden gemodelleerd. Voor hydratatiewater vertonen de bijdragen van translatie- en rotatiebewegingen van watermoleculen een andere afhankelijkheid van de momentumoverdracht q, wat hun onderscheid in het data-analyseproces mogelijk maakt17.
Dit artikel illustreert de neutronen-backscatering-methode met de studie van eiwitten die zich bleken te kunnen ontvouwen, aggregeren tot een canonieke vorm bestaande uit stapels β-strengen – het zogenaamde cross-β-patroon19,20 – en langwerpige vezels vormen. Dit is de zogenaamde amyloïde aggregatie, die uitgebreid wordt bestudeerd vanwege zijn centrale rol bij neurodegeneratieve aandoeningen zoals de ziekte van Alzheimer of Parkinson21,22. De studie van de amyloïde eiwitten wordt ook gemotiveerd door de functionele rol die ze kunnen spelen 23,24 of hun hoge potentieel voor de ontwikkeling van nieuwe biomaterialen25. De fysisch-chemische determinanten van de amyloïde aggregatie blijven onduidelijk en er is geen algemene theorie van amyloïde aggregatie beschikbaar, ondanks enorme vooruitgang in de afgelopen jaren21,26.
Amyloïde aggregatie impliceert veranderingen in eiwitstructuur en stabiliteit met de tijd, waarvan de studie van nature dynamiek impliceert, gekoppeld aan eiwitconformatiestabiliteit, eiwitfunctie en eiwitenergielandschap27. Dynamica is direct gekoppeld aan de stabiliteit van een specifieke toestand door de entropische bijdrage voor de snelste bewegingen28, en de eiwitfunctie kan worden ondersteund door bewegingen op verschillende tijdschalen van sub-ps voor lichtgevoelige eiwitten29 tot ms voor domeinbewegingen, die kunnen worden vergemakkelijkt door picoseconde-nanoseconde dynamica30.
Twee voorbeelden van het gebruik van neutronenbackscattering spectroscopie om amyloïde eiwitten te bestuderen zullen worden gepresenteerd, één in vloeibare toestand om eiwitdynamica te bestuderen en één in gehydrateerde poedertoestand om hydratatiewaterdynamica te bestuderen. Het eerste voorbeeld betreft de aggregatie van lysozym in bolletjes ter grootte van μm (deeltjes genoemd) gevolgd in real time5, en het tweede een vergelijking van de waterdynamiek in inheemse en geaggregeerde toestanden van het menselijke eiwit tau31.
Lysozym is een enzym dat betrokken is bij de immuunafweer en bestaat uit 129 aminozuurresiduen. Lysozym kan deeltjes vormen in gedeutereerde buffer bij pD van 10,5 en bij een temperatuur van 90 °C. Met neutronenverstrooiing toonden we aan dat de tijdsevolutie van de diffusiecoëfficiënt van het massamiddelpunt van lysozym de enkele exponentiële kinetiek van thioflavine-T-fluorescentie volgt (een fluorescerende sonde die wordt gebruikt om de vorming van amyloïde cross-β-patronen32 te volgen, wat aangeeft dat de vorming van deeltjessuperstructuren en kruis-β patronen in één stap met dezelfde snelheid plaatsvinden. Bovendien bleef de interne dynamiek constant gedurende het aggregatieproces, wat kan worden verklaard door een snelle conformatieverandering die niet kan worden waargenomen op NBS-instrumenten, of door de afwezigheid van significante verandering in interne eiwitenergie bij aggregatie.
Het menselijke eiwit tau is een intrinsiek ongeordend eiwit (IDP) bestaande uit 441 aminozuren voor de zogenaamde 2N4R-isovorm, die met name betrokken is bij de ziekte van Alzheimer33. Met behulp van neutronenverstrooiing op poeders van geperdeutereerd eiwit tau, toonden we aan dat de hydratatiewaterdynamiek wordt verhoogd in de vezeltoestand, met een hogere populatie watermoleculen die translationele bewegingen ondergaan. Het resultaat suggereert dat een toename van hydratatiewaterentropie de amyloïde fibrillatie van tau zou kunnen veroorzaken.
