Das Neutron ist ein ladungsloses und massereiches Teilchen, das im Laufe der Jahre erfolgreich zur Untersuchung von Proben in verschiedenen Bereichen von der Grundlagenphysik bis zur Biologieeingesetzt wurde 1. Für biologische Anwendungen werden Kleinwinkel-Neutronenstreuung, inelastische Neutronenstreuung sowie Neutronenkristallographie und Reflektometrie häufig eingesetzt 2,3,4. Die inelastische Neutronenstreuung ermöglicht eine ensemblegemittelte Messung der Dynamik, ohne dass eine spezifische Markierung an sich erforderlich ist, und eine Signalqualität, die nicht von der Größe oder dem Protein abhängt5. Die Messung kann in einer hochkomplexen Umgebung für das untersuchte Protein durchgeführt werden, die das intrazelluläre Medium nachahmt, wie z. B. ein deuteriertes bakterielles Lysat oder sogar in vivo 3,6,7. Verschiedene experimentelle Aufbauten können verwendet werden, um die Dynamik zu untersuchen, nämlich i) Flugzeit-gebender Zugang zu sub-ps-ps-Dynamik, ii) Rückstreuung-gebender Zugang zu ps-ns-Dynamik und iii) Spin-Echo-gebender Zugang zu Dynamik von ns bis zu Hunderten von ns. Die Neutronenrückstreuung nutzt das Braggsche Gesetz 2d sinθ = nλ, wobei d der Abstand zwischen Ebenen in einem Kristall, θ der Streuwinkel, n die Streuordnung und λ die Wellenlänge ist. Die Verwendung von Kristallen für die Rückstreuung zu den Detektoren ermöglicht es, eine hohe Energieauflösung von typischerweise ~0,8 μeV zu erreichen. Um den Energieaustausch zu messen, wird entweder ein Dopplerantrieb verwendet, der einen Kristall in der Rückstreuung trägt, um die einfallende Neutronenwellenlänge 8,9,10 zu definieren und einzustellen (Abbildung 1), oder es kann ein Time-of-Flight-Setup auf Kosten einer Verringerung der Energieauflösung 11 verwendet werden.
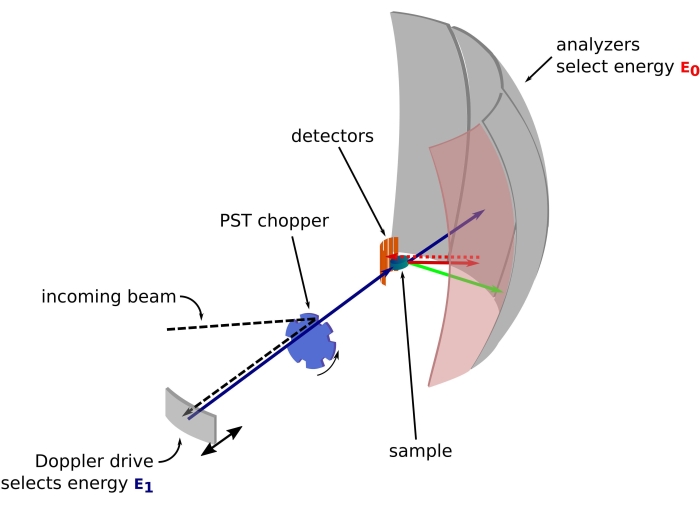
Abbildung 1: Skizze eines Neutronenrückstreuspektrometers mit Dopplerantrieb. Der einfallende Strahl trifft auf den Phasenraumtransformations-Chopper42 (PST), der den Fluss an der Probenposition erhöht. Es wird dann durch den Dopplerantrieb, der eine Energie E1 (cyanfarbener Pfeil) wählt, in Richtung der Probe zurückgestreut. Die Neutronen werden dann von der Probe gestreut (mit unterschiedlichen Energien, die durch die Farbe der Pfeile dargestellt werden) und die Analysatoren, die aus Si 111-Kristallen bestehen, streuen nur Neutronen mit einer spezifischen Energie E0 zurück (hier rote Pfeile). Daher wird der Impulstransfer q aus der detektierten Position des Neutrons auf dem Detektorarray und der Energietransfer aus der Differenz E1 – E0 erhalten. Die erwartete Flugzeit für den vom PST erzeugten Neutronenpuls wird verwendet, um das Signal der Neutronen zu verwerfen, die direkt in Richtung der Detektorröhren gestreut werden. Abkürzung: PST = Phasenraumtransformation. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
Für die Rückstreuspektroskopie kommt der Hauptbeitrag zum Signal von protonenreichen Wasserstoffproben, wie z. B. Proteinen, aus der inkohärenten Streuung, für die die Streuintensität Sinc(q, ω) durch Gl (1)12 gezeigt wird
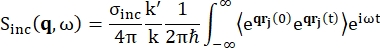 (1)
(1)
Dabei ist σinc der inkohärente Wirkungsquerschnitt des betrachteten Elements, k’ die Norm des gestreuten Wellenvektors, k die Norm des eingehenden Wellenvektors, q (= k – k’) die Impulsübertragung, r j(t) der Ortsvektor des Atoms j zum Zeitpunkt t und ω die Frequenz, die dem Energietransfer zwischen dem einfallenden Neutron und dem System entspricht. Die eckigen Klammern bezeichnen den Ensembledurchschnitt. Die inkohärente Streuung untersucht daher die ensemblegemittelte Einzelteilchen-Selbstkorrelation der Atompositionen mit der Zeit und liefert die Selbstdynamik gemittelt über alle Atome im System und verschiedene Zeitursprünge (Ensemble-Mittelwert). Die Streufunktion ist die zeitliche Fourier-Transformation der intermediären Streufunktion I(q, t), die als Fourier-Transformation im Raum der van-Hove-Korrelationsfunktion angesehen werden kann, die durch Gl (2) gezeigt wird:
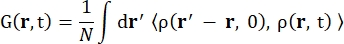 (2)
(2)
Wobei ρ(r,t) die Wahrscheinlichkeitsdichte ist, ein Atom an der Position r und der Zeit t 13 zu finden.
Für einen Fickschen Diffusionsprozess ergibt sich die Selbstdiffusionsfunktion (siehe Gl (3)) nach einer doppelten Fourier-Transformation in einer Streufunktion, die aus einem Lorentz-Operator mit Linienbreite besteht, der durch γ = Dq2 gegeben ist.
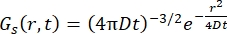 (3)
(3)
Ausgefeiltere Modelle wurden entwickelt und für nützlich befunden, wie z.B. das Sprungdiffusionsmodell von Singwi und Sjölander für die interne ps-ns-Proteindynamik14 oder das Rotationsmodell von Sears für Hydratationswasser15,16,17.
Auf dem Neutronenrückstreuinstrument IN16B 8,9 am ILL in Grenoble, Frankreich (Supplemental Figure S1), besteht ein bei Proteinen üblicherweise verwendeter Aufbau aus Si 111-Kristallen für die Analysatoren mit einem Dopplerantrieb zur Abstimmung der eingehenden Wellenlänge (Supplemental Figure S2A), wodurch der Impulsübertragungsbereich ~0,2 Å-1 < q < ~2 Å-1 und der Energieübertragungsbereich von –30 μeV < 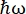 < 30 μeV, was Zeitskalen von wenigen ps bis einigen ns und Entfernungen von einigen Å entspricht. Darüber hinaus bietet IN16B die Möglichkeit, elastische und inelastische Fixed-Window-Scans (E/IFWS)10 durchzuführen, die die Datenerfassung bei einer festen Energieübertragung beinhalten. Da der Fluss bei der Arbeit mit Neutronen begrenzt ist, ermöglicht E/IFWS die Maximierung des Flusses für einen Energietransfer, wodurch die Erfassungszeit reduziert wird, die benötigt wird, um ein zufriedenstellendes Signal-Rausch-Verhältnis zu erhalten. Eine neuere Option ist das Rückstreu- und Flugzeitspektrometer (BATS) Mode11, das die Messung eines breiten Bereichs von Energietransfers (z. B. -150 μeV < < 150 μeV) mit einem höheren Fluss als mit dem Dopplerantrieb, jedoch auf Kosten einer geringeren Energieauflösung ermöglicht (ergänzende Abbildung S2B).
< 30 μeV, was Zeitskalen von wenigen ps bis einigen ns und Entfernungen von einigen Å entspricht. Darüber hinaus bietet IN16B die Möglichkeit, elastische und inelastische Fixed-Window-Scans (E/IFWS)10 durchzuführen, die die Datenerfassung bei einer festen Energieübertragung beinhalten. Da der Fluss bei der Arbeit mit Neutronen begrenzt ist, ermöglicht E/IFWS die Maximierung des Flusses für einen Energietransfer, wodurch die Erfassungszeit reduziert wird, die benötigt wird, um ein zufriedenstellendes Signal-Rausch-Verhältnis zu erhalten. Eine neuere Option ist das Rückstreu- und Flugzeitspektrometer (BATS) Mode11, das die Messung eines breiten Bereichs von Energietransfers (z. B. -150 μeV < < 150 μeV) mit einem höheren Fluss als mit dem Dopplerantrieb, jedoch auf Kosten einer geringeren Energieauflösung ermöglicht (ergänzende Abbildung S2B).
Eine wichtige Eigenschaft der Neutronenstreuung ist, dass der inkohärente Wirkungsquerschnitt σinc für Wasserstoff einen 40-mal höheren Wert als für Deuterium hat und für andere Elemente, die üblicherweise in biologischen Proben vorkommen, vernachlässigbar ist. Daher kann die Dynamik von Proteinen in einer flüssigen Umgebung unter Verwendung eines deuterierten Puffers untersucht werden, und der Pulverzustand ermöglicht die Untersuchung entweder der inneren Dynamik von Proteinen mit hydriertem Proteinpulver, das mit D2O hydratisiert ist, oder die Untersuchung von Hydratationswasser für perdeuteriertes Proteinpulver, das mitH2Ohydratisiert ist. Im flüssigen Zustand ermöglicht die Neutronenrückstreuung typischerweise den gleichzeitigen Zugriff auf die Selbstdiffusion des Massenschwerpunkts von Proteinen (Fickian-Typ-Diffusion) und deren interne Dynamik. Bei letzteren handelt es sich um Rückgrat- und Seitenkettenbewegungen, die in der Regel durch das sogenannte Sprungdiffusionsmodell oder andere beschrieben werden 3,18. In hydrierten Proteinpulvern fehlt die Proteindiffusion und es muss nur die interne Dynamik modelliert werden. Für Hydratationswasser weisen die Beiträge von Translations- und Rotationsbewegungen von Wassermolekülen eine andere Abhängigkeit vom Impulstransfer q auf, was ihre Unterscheidung im Datenanalyseprozess ermöglicht17.
Diese Arbeit veranschaulicht die Methode der Neutronenrückstreuung anhand der Untersuchung von Proteinen, die in der Lage sind, sich zu entfalten, zu einer kanonischen Form zu aggregieren, die aus Stapeln von β-Strängen besteht – dem sogenannten Kreuz-β-Muster19,20 – und längliche Fasern zu bilden. Dabei handelt es sich um die sogenannte Amyloid-Aggregation, die aufgrund ihrer zentralen Rolle bei neurodegenerativen Erkrankungen wie Alzheimer oder Parkinson ausgiebig untersucht wird21,22. Die Untersuchung der Amyloid-Proteine ist auch durch die funktionelle Rolle, die sie spielen können, motiviert 23,24 oder durch ihr hohes Potenzial für die Entwicklung neuartiger Biomaterialien25. Die physikalisch-chemischen Determinanten der Amyloid-Aggregation sind nach wie vor unklar, und es gibt keine allgemeine Theorie der Amyloid-Aggregation, trotz enormer Fortschritte in den letzten Jahren21,26.
Die Amyloid-Aggregation impliziert Veränderungen in der Proteinstruktur und -stabilität im Laufe der Zeit, deren Untersuchung natürlich eine Dynamik impliziert, die mit der Stabilität der Proteinkonformation, der Proteinfunktion und der Proteinenergielandschaft zusammenhängt27. Die Dynamik ist durch den entropischen Beitrag für die schnellsten Bewegungen28 direkt mit der Stabilität eines bestimmten Zustands verbunden, und die Proteinfunktion kann durch Bewegungen auf verschiedenen Zeitskalen von sub-ps für lichtempfindliche Proteine29 bis ms für Domänenbewegungen aufrechterhalten werden, was durch die Pikosekunden-Nanosekunden-Dynamik30 erleichtert werden kann.
Es werden zwei Beispiele für die Verwendung der Neutronenrückstreuspektroskopie zur Untersuchung von Amyloidproteinen vorgestellt, eines im flüssigen Zustand, um die Proteindynamik zu untersuchen, und eines im hydratisierten Pulverzustand, um die Dynamik des Hydratationswassers zu untersuchen. Das erste Beispiel betrifft die Aggregation von Lysozym in μm-große Kügelchen (Partikel genannt), gefolgt in Echtzeit5, und das zweite ein Vergleich der Wasserdynamik in nativen und aggregierten Zuständen des menschlichen Proteins Tau31.
Lysozym ist ein Enzym, das an der Immunabwehr beteiligt ist und aus 129 Aminosäureresten besteht. Lysozym kann in deuteriertem Puffer bei einem pD-Wert von 10,5 und einer Temperatur von 90 °C Partikel bilden. Mit Hilfe der Neutronenstreuung konnten wir zeigen, dass die zeitliche Entwicklung des Massenschwerpunkt-Diffusionskoeffizienten von Lysozym der einzelnen exponentiellen Kinetik der Thioflavin-T-Fluoreszenz folgt (eine Fluoreszenzsonde, die zur Überwachung der Bildung von Amyloid-Kreuz-β-Musternverwendet wird 32), was darauf hindeutet, dass die Bildung von partikulären Überstrukturen und Kreuz-β-Mustern in einem einzigen Schritt mit der gleichen Geschwindigkeit erfolgt. Darüber hinaus blieb die interne Dynamik während des gesamten Aggregationsprozesses konstant, was entweder durch eine schnelle Konformationsänderung erklärt werden kann, die mit NBS-Instrumenten nicht beobachtet werden kann, oder durch das Fehlen einer signifikanten Änderung der internen Proteinenergie bei der Aggregation.
Das menschliche Protein Tau ist ein intrinsisch ungeordnetes Protein (IDP), das aus 441 Aminosäuren für die sogenannte 2N4R-Isoform besteht, die insbesondere an der Alzheimer-Krankheit beteiligt ist33. Mit Hilfe der Neutronenrückstreuung an Pulvern aus perdeuteriertem Proteintau konnten wir zeigen, dass die Dynamik des Hydratationswassers im Faserzustand erhöht ist, wobei eine höhere Population von Wassermolekülen Translationsbewegungen durchläuft. Das Ergebnis deutet darauf hin, dass eine Erhöhung der Entropie des Hydratationswassers das Amyloid-Flimmern von Tau antreiben könnte.
