Il neutrone è una particella massiccia e priva di carica che è stata utilizzata con successo nel corso degli anni per sondare campioni in vari campi, dalla fisica fondamentale alla biologia1. Per le applicazioni biologiche, lo scattering neutronico a piccolo angolo, lo scattering neutronico anelastico, la cristallografia neutronica e la riflettometria sono ampiamente utilizzati 2,3,4. Lo scattering anelastico di neutroni fornisce una misura mediata dell’insieme della dinamica senza richiedere un’etichettatura specifica di per sé e una qualità del segnale che non dipende dalla dimensione o dalla proteina5. La misurazione può essere effettuata utilizzando un ambiente altamente complesso per la proteina in studio che imita il mezzo intracellulare, come un lisato batterico deuterato o anche in vivo 3,6,7. Diverse configurazioni sperimentali possono essere utilizzate per studiare le dinamiche, vale a dire i) l’accesso al tempo di volo alle dinamiche sub-ps-ps, ii) l’accesso alla dinamica ps-ns e iii) l’accesso alle dinamiche da ns a centinaia di ns. La retrodiffusione neutronica fa uso della legge di Bragg 2d sinθ = nλ, dove d è la distanza tra i piani in un cristallo, θ l’angolo di scattering, n l’ordine di scattering e λ la lunghezza d’onda. L’uso di cristalli per il backscattering verso i rivelatori consente di ottenere un’alta risoluzione in energia, tipicamente ~ 0,8 μeV. Per misurare lo scambio di energia, viene utilizzato un azionamento Doppler che trasporta un cristallo in retrodiffusione per definire e sintonizzare la lunghezza d’onda dei neutroni in arrivo 8,9,10 (Figura 1), oppure è possibile utilizzare una configurazione del tempo di volo al costo di una diminuzione della risoluzione energetica 11.
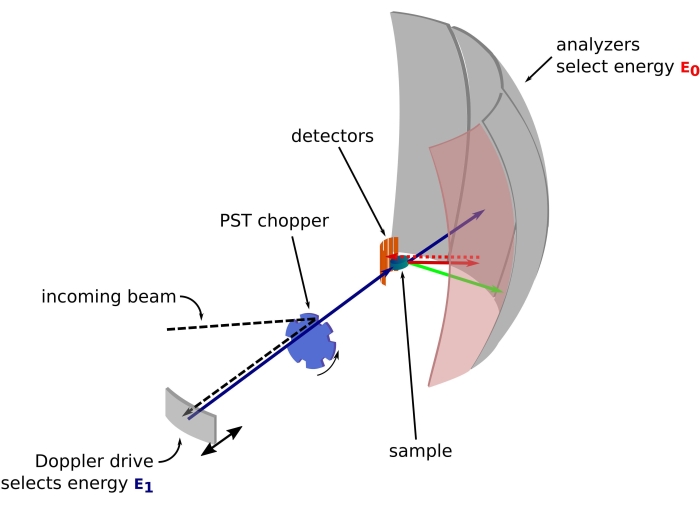
Figura 1: Schizzo di uno spettrometro di retrodiffusione di neutroni con un drive Doppler. Il fascio in entrata colpisce il chopper PST (Phase Space Transformation)42, che aumenta il flusso nella posizione del campione. Viene quindi retrodiffuso verso il campione dall’azionamento Doppler, che seleziona un’energia E1 (freccia ciano). I neutroni vengono poi dispersi dal campione (con energie diverse rappresentate dal colore delle frecce) e gli analizzatori, costituiti da cristalli di Si 111, disperderanno solo neutroni con una specifica energia E0 (qui frecce colorate di rosso). Quindi, il trasferimento di quantità di moto q è ottenuto dalla posizione rilevata del neutrone sull’array del rivelatore e il trasferimento di energia è ottenuto dalla differenza E1– E0. Il tempo di volo previsto per l’impulso neutronico prodotto dal PST viene utilizzato per scartare il segnale dai neutroni sparsi direttamente verso i tubi del rivelatore. Abbreviazione: PST = trasformazione dello spazio delle fasi. Fare clic qui per visualizzare una versione ingrandita di questa figura.
Per la spettroscopia di retrodiffusione, il principale contributo al segnale da campioni ricchi di protoni di idrogeno, come le proteine, proviene dallo scattering incoerente, per il quale l’intensità di scattering Sinc(q, ω) è mostrata da Eq (1)12
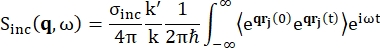 (1)
(1)
Dove σinc è la sezione trasversale incoerente dell’elemento considerato, k’ è la norma del vettore d’onda diffuso, k la norma del vettore d’onda entrante, q (= k – k’) il trasferimento di quantità di moto, r j(t) il vettore posizione dell’atomo j al tempo t e ω la frequenza corrispondente al trasferimento di energia tra il neutrone entrante e il sistema. Le parentesi angolari indicano la media dell’ensemble. Quindi, lo scattering incoerente sonda l’autocorrelazione delle singole particelle mediata dall’insieme delle posizioni degli atomi con il tempo e fornisce l’autodinamica mediata su tutti gli atomi nel sistema e le diverse origini temporali (media dell’insieme). La funzione di scattering è la trasformata di Fourier nel tempo della funzione di scattering intermedia I(q, t), che può essere vista come la trasformata di Fourier nello spazio della funzione di correlazione di van Hove mostrata da Eq (2):
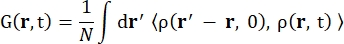 (2)
(2)
Dove ρ(r,t) è la densità di probabilità di trovare un atomo in posizione r e tempo t 13.
Per un processo di diffusione di Fick, la funzione di autodiffusione risulta (vedi Eq (3)) dopo una doppia trasformata di Fourier in una funzione di scattering consistente in un lorentziano di larghezza di linea data da γ = Dq2.
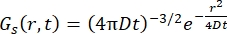 (3)
(3)
Sono stati sviluppati e trovati utili modelli più sofisticati come il modello di diffusione a salto di Singwi e Sjölander per la dinamica interna delle proteine ps-ns14 o il modello di rotazione di Sears per l’acqua di idratazione15,16,17.
Sullo strumento di retrodiffusione neutronica (NBS) IN16B 8,9 presso l’ILL, Grenoble, Francia (Figura supplementare S1), una configurazione comunemente usata con le proteine è costituita da cristalli di Si 111 per gli analizzatori con un drive Doppler per la regolazione della lunghezza d’onda in entrata (Figura supplementare S2A), dando così accesso all’intervallo di trasferimento della quantità di moto ~0,2 Å-1 < q < ~2 Å-1 e intervallo di trasferimento di energia di –30 μeV < 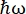 < 30 μeV, corrispondenti a scale temporali che vanno da pochi ps a pochi ns e distanze di pochi Å. Inoltre, IN16B offre la possibilità di eseguire scansioni elastiche e anelastiche a finestra fissa (E/IFWS)10, che includono l’acquisizione dei dati a un trasferimento di energia fisso. Poiché il flusso è limitato quando si lavora con neutroni, E/IFWS consente di massimizzare il flusso per un trasferimento di energia, riducendo così il tempo di acquisizione necessario per ottenere un soddisfacente rapporto segnale-rumore. Un’opzione più recente è il backscattering and time-of-flight spectrometer (BATS) mode11, che consente la misurazione di un’ampia gamma di trasferimenti di energia, (ad esempio, -150 μeV < < 150 μeV), con un flusso più elevato rispetto al Doppler, ma al costo di una risoluzione energetica inferiore (Figura supplementare S2B).
< 30 μeV, corrispondenti a scale temporali che vanno da pochi ps a pochi ns e distanze di pochi Å. Inoltre, IN16B offre la possibilità di eseguire scansioni elastiche e anelastiche a finestra fissa (E/IFWS)10, che includono l’acquisizione dei dati a un trasferimento di energia fisso. Poiché il flusso è limitato quando si lavora con neutroni, E/IFWS consente di massimizzare il flusso per un trasferimento di energia, riducendo così il tempo di acquisizione necessario per ottenere un soddisfacente rapporto segnale-rumore. Un’opzione più recente è il backscattering and time-of-flight spectrometer (BATS) mode11, che consente la misurazione di un’ampia gamma di trasferimenti di energia, (ad esempio, -150 μeV < < 150 μeV), con un flusso più elevato rispetto al Doppler, ma al costo di una risoluzione energetica inferiore (Figura supplementare S2B).
Una proprietà importante dello scattering neutronico è che la sezione d’urto incoerente σinc ha un valore 40 volte superiore per l’idrogeno che per il deuterio ed è trascurabile per altri elementi che si trovano comunemente nei campioni biologici. Pertanto, la dinamica delle proteine in un ambiente liquido può essere studiata utilizzando un tampone deuterato e lo stato della polvere consente lo studio della dinamica interna delle proteine con polvere proteica idrogenata idratata conD 2 O, o lo studio dell’acqua di idratazione per proteine perdeuterate in polvere idratate con H2O. Allo stato liquido, la retrodiffusione neutronica consente tipicamente l’accesso simultaneo all’autodiffusione del centro di massa delle proteine (diffusione di tipo Fickiano) e alla loro dinamica interna. Questi ultimi sono movimenti della spina dorsale e della catena laterale solitamente descritti dal cosiddetto modello di diffusione del salto o altri 3,18. Nelle polveri proteiche idrogenate, la diffusione proteica è assente e deve essere modellata solo la dinamica interna. Per l’acqua di idratazione, i contributi dei moti traslazionali e rotazionali delle molecole d’acqua presentano una diversa dipendenza dal trasferimento di quantità di moto q, che consente la loro distinzione nel processo di analisi dei dati17.
Questo articolo illustra il metodo di retrodiffusione neutronica con lo studio di proteine che sono risultate in grado di dispiegarsi, aggregarsi in una forma canonica costituita da pile di β-filamenti – il cosiddetto modello di β incrociato19,20 – e formare fibre allungate. Si tratta della cosiddetta aggregazione amiloide, ampiamente studiata per il suo ruolo centrale nelle malattie neurodegenerative come il morbo di Alzheimer o il Parkinson21,22. Lo studio delle proteine amiloidi è anche motivato dal ruolo funzionale che possono svolgere 23,24 o dal loro elevato potenziale per lo sviluppo di nuovi biomateriali25. I determinanti fisico-chimici dell’aggregazione amiloide rimangono poco chiari e non è disponibile alcuna teoria generale dell’aggregazione amiloide, nonostante gli enormi progressi compiuti negli ultimi anni21,26.
L’aggregazione amiloide implica cambiamenti nella struttura e nella stabilità delle proteine nel tempo, il cui studio implica naturalmente dinamiche, legate alla stabilità della conformazione delle proteine, alla funzione proteica e al panorama energetico delle proteine27. La dinamica è direttamente collegata alla stabilità di uno stato specifico attraverso il contributo entropico per i moti più veloci28, e la funzione proteica può essere sostenuta da movimenti su varie scale temporali da sub-ps per proteine sensibili alla luce29 a ms per movimenti di dominio, che possono essere facilitati dalla dinamica picosecondo-nanosecondo30.
Saranno presentati due esempi di utilizzo della spettroscopia di retrodiffusione neutronica per studiare le proteine amiloidi, uno allo stato liquido per studiare la dinamica delle proteine e uno allo stato di polvere idratata per studiare la dinamica dell’acqua di idratazione. Il primo esempio riguarda l’aggregazione del lisozima in sfere di dimensioni μm (chiamate particolato) seguite in tempo reale5, e il secondo un confronto della dinamica dell’acqua negli stati nativi e aggregati della proteina umana tau31.
Il lisozima è un enzima coinvolto nella difesa immunitaria ed è composto da 129 residui di aminoacidi. Il lisozima può formare particolato in tampone deuterato a pD di 10,5 e ad una temperatura di 90 °C. Con lo scattering neutronico, abbiamo dimostrato che l’evoluzione temporale del coefficiente di diffusione del centro di massa del lisozima segue la singola cinetica esponenziale della fluorescenza T della tioflavina (una sonda fluorescente utilizzata per monitorare la formazione di modelli di β incrociato amiloide32), indicando che le sovrastrutture particolate di formazione e i modelli di cross-β si verificano in un unico passaggio con la stessa velocità. Inoltre, la dinamica interna è rimasta costante durante tutto il processo di aggregazione, che può essere spiegato sia da un rapido cambiamento conformazionale che non può essere osservato sugli strumenti NBS, sia dall’assenza di cambiamenti significativi nell’energia interna della proteina dopo l’aggregazione.
La proteina umana tau è una proteina intrinsecamente disordinata (IDP) costituita da 441 aminoacidi per la cosiddetta isoforma 2N4R, che è particolarmente coinvolta nella malattia di Alzheimer33. Utilizzando la retrodiffusione neutronica su polveri di proteina tau perdeuterata, abbiamo dimostrato che la dinamica dell’acqua di idratazione è aumentata allo stato di fibra, con una maggiore popolazione di molecole d’acqua sottoposte a movimenti traslazionali. Il risultato suggerisce che un aumento dell’entropia dell’acqua di idratazione potrebbe guidare la fibrillazione amiloide della tau.
