Nøytronet er en ladningsløs og massiv partikkel som har blitt brukt gjennom årene til å undersøke prøver på ulike felt fra grunnleggende fysikk til biologi1. For biologiske anvendelser er småvinklet nøytronspredning, uelastisk nøytronspredning og nøytronkrystallografi og reflektometri mye brukt 2,3,4. Uelastisk nøytronspredning gir en ensemble-gjennomsnittlig måling av dynamikken uten å kreve spesifikk merking i seg selv, og en signalkvalitet som ikke avhenger av størrelsen eller proteinet5. Målingen kan gjøres ved hjelp av et svært komplekst miljø for proteinet som studeres som etterligner det intracellulære mediet, for eksempel et deuterert bakterielt lysat eller til og med in vivo 3,6,7. Ulike eksperimentelle oppsett kan brukes til å studere dynamikken, nemlig i) time-of-flight-gi tilgang til sub-ps-ps-dynamikk, ii) backscattering-gi tilgang til ps-ns dynamikk, og iii) spin-echo-gi tilgang til dynamikk fra ns til hundrevis av ns. Nøytron-backscattering gjør bruk av Braggs lov 2d sinθ = nλ, hvor d er avstanden mellom plan i en krystall, θ spredningsvinkelen, n spredningsrekkefølgen og λ bølgelengden. Bruken av krystaller for tilbakespredning mot detektorene gjør det mulig å oppnå en høy oppløsning i energi, typisk ~0,8 μeV. For å måle energiutvekslingen brukes enten en Doppler-stasjon som bærer en krystall i tilbakespredning til å definere og justere den innkommende nøytronbølgelengden 8,9,10 (figur 1), eller et flytidsoppsett kan brukes på bekostning av en reduksjon i energioppløsning 11.
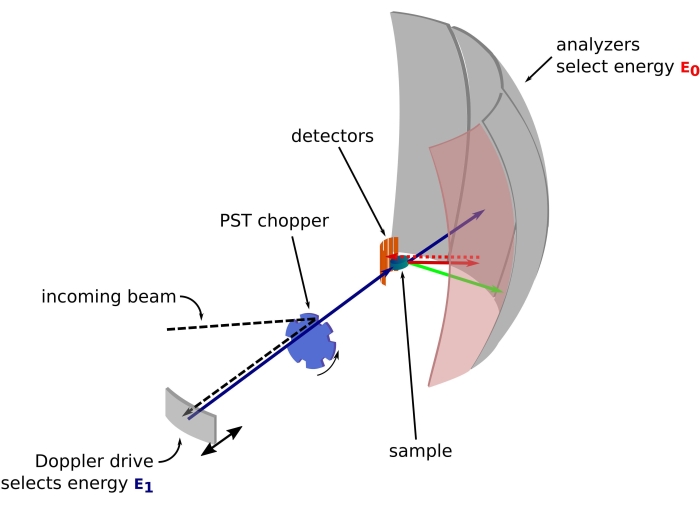
Figur 1: Skisse av et nøytron-tilbakespredningsspektrometer med dopplerdrift. Den innkommende strålen treffer faseromtransformasjonen (PST) helikopter42, noe som øker fluksen ved prøveposisjonen. Den blir deretter tilbakespredt mot prøven av Doppler-stasjonen, som velger en energi E1 (cyan pil). Nøytronene blir deretter spredt av prøven (med forskjellige energier representert ved pilens farge) og analysatorene, laget av Si 111-krystaller, vil bare tilbakespre nøytroner med en spesifikk energi E0 (rødfargede piler her). Derfor oppnås momentumoverføringen q fra den oppdagede posisjonen til nøytronet på detektormatrisen, og energioverføringen oppnås fra forskjellen E1– E0. Flytiden som forventes for nøytronpulsen produsert av PST, brukes til å forkaste signalet fra nøytronene spredt direkte mot detektorrørene. Forkortelse: PST = faseromtransformasjon. Klikk her for å se en større versjon av denne figuren.
For tilbakespredningsspektroskopi kommer hovedbidraget til signalet fra hydrogenprotonrike prøver, slik som proteiner, fra usammenhengende spredning, for hvilken spredningsintensiteten Sinc(q, ω) er vist ved Eq (1)12
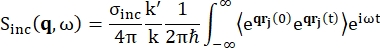 (1)
(1)
Hvor σinc er det usammenhengende tverrsnittet av elementet som vurderes, er k’ normen for den spredte bølgevektoren, k normen for den innkommende bølgevektoren, q (= k – k’) momentumoverføringen, r j (t) posisjonsvektoren til atom j på tidspunkt t, og ω frekvensen som tilsvarer energioverføringen mellom det innkommende nøytronet og systemet. Vinkelparentesene angir ensemblegjennomsnittet. Derfor sonderer inkoherent spredning ensemble-gjennomsnittlig enkeltpartikkel selvkorrelasjon av atomposisjoner med tiden og gir selvdynamikken gjennomsnittet over alle atomer i systemet og forskjellige tidsopprinnelser (ensemblegjennomsnitt). Spredningsfunksjonen er Fouriertransformasjonen i tid for den mellomliggende spredningsfunksjonen I (q, t), som kan sees på som Fourier-transformasjonen i rommet til van Hove-korrelasjonsfunksjonen vist ved Eq (2):
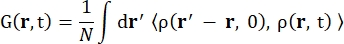 (2)
(2)
Hvor ρ(r,t) er sannsynlighetstettheten for å finne et atom i posisjon r og tid t 13.
For en Fickian diffusjonsprosess resulterer selvdiffusjonsfunksjonen (se Eq (3)) etter en dobbel Fourier-transformasjon i en spredningsfunksjon bestående av en Lorentzian med linjebredde gitt ved γ = Dq2.
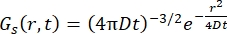 (3)
(3)
Mer sofistikerte modeller ble utviklet og funnet nyttige, for eksempel hoppdiffusjonsmodellen av Singwi og Sjölander for ps-ns intern proteindynamikk14 eller rotasjonsmodellen av Sears for hydreringsvann15,16,17.
På nøytronbackscattering-instrumentet (NBS) IN16B8,9 ved ILL, Grenoble, Frankrike (tilleggsfigur S1), består et oppsett som vanligvis brukes med proteiner av Si 111-krystaller for analysatorene med en Doppler-stasjon for innstilling av den innkommende bølgelengden (tilleggsfigur S2A), og gir dermed tilgang til momentumoverføringsområdet ~0,2 Å-1 < q < ~2 Å-1 og energioverføringsområde på –30 μeV < 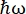 < 30 μeV-tilsvarende tidsskalaer fra noen få ps til noen få ns og avstander på noen få Å. I tillegg tilbyr IN16B muligheten til å utføre elastiske og uelastiske skanninger med faste vinduer (E/IFWS)10, som inkluderer datainnsamling ved en fast energioverføring. Siden fluksen er begrenset ved arbeid med nøytroner, tillater E / IFWS maksimering av fluksen for en energioverføring, og reduserer dermed innsamlingstiden som trengs for å oppnå et tilfredsstillende signal-støyforhold. Et nyere alternativ er backscattering and time-of-flight spectrometer (BATS) modus11, som tillater måling av et bredt spekter av energioverføringer, (f.eks. -150 μeV < < 150 μeV), med en høyere fluks enn med Doppler-stasjonen, men på bekostning av en lavere energioppløsning (tilleggsfigur S2B).
< 30 μeV-tilsvarende tidsskalaer fra noen få ps til noen få ns og avstander på noen få Å. I tillegg tilbyr IN16B muligheten til å utføre elastiske og uelastiske skanninger med faste vinduer (E/IFWS)10, som inkluderer datainnsamling ved en fast energioverføring. Siden fluksen er begrenset ved arbeid med nøytroner, tillater E / IFWS maksimering av fluksen for en energioverføring, og reduserer dermed innsamlingstiden som trengs for å oppnå et tilfredsstillende signal-støyforhold. Et nyere alternativ er backscattering and time-of-flight spectrometer (BATS) modus11, som tillater måling av et bredt spekter av energioverføringer, (f.eks. -150 μeV < < 150 μeV), med en høyere fluks enn med Doppler-stasjonen, men på bekostning av en lavere energioppløsning (tilleggsfigur S2B).
En viktig egenskap ved nøytronspredning er at det usammenhengende tverrsnittet σinc har en 40 ganger høyere verdi for hydrogen enn for deuterium og er ubetydelig for andre elementer som ofte finnes i biologiske prøver. Derfor kan dynamikken til proteiner i et flytende miljø studeres ved å bruke en deuterert buffer, og pulvertilstanden tillater studier av enten proteinintern dynamikk med hydrogenert proteinpulver hydrert med D 2 O, eller studiet av hydreringsvann for perdeuterert proteinpulver hydrert med H2O. I flytende tilstand tillater nøytron-backscattering vanligvis samtidig tilgang til massesenteret selvdiffusjon av proteiner (Fickian-type diffusjon) og deres indre dynamikk. Sistnevnte er ryggrads- og sidekjedebevegelser som vanligvis beskrives av den såkalte hoppdiffusjonsmodellen eller andre 3,18. I hydrogenerte proteinpulver er proteindiffusjonen fraværende, og bare intern dynamikk må modelleres. For hydreringsvann presenterer bidragene fra translasjons- og rotasjonsbevegelser av vannmolekyler en annen avhengighet av momentumoverføringen q, noe som gjør det mulig å skille dem i dataanalyseprosessen17.
Dette papiret illustrerer nøytron-backscattering-metoden med studiet av proteiner som ble funnet å kunne utfolde seg, aggregere til en kanonisk form bestående av stabler av β-tråder – det såkalte kryss-β mønsteret19,20 – og danne langstrakte fibre. Dette er den såkalte amyloidaggregeringen, som er grundig studert på grunn av sin sentrale rolle i nevrodegenerative lidelser som Alzheimers eller Parkinsons sykdommer21,22. Studien av amyloidproteinene er også motivert av den funksjonelle rollen de kan spille 23,24 eller deres høye potensial for utvikling av nye biomaterialer25. De fysisk-kjemiske determinantene for amyloidaggregeringen er fortsatt uklare, og ingen generell teori om amyloidaggregering er tilgjengelig, til tross for enorm fremgang de siste årene21,26.
Amyloidaggregering innebærer endringer i proteinstruktur og stabilitet med tiden, hvis studie naturlig innebærer dynamikk, knyttet til proteinkonformasjonsstabilitet, proteinfunksjon og proteinenergilandskap27. Dynamikk er direkte knyttet til stabiliteten til en bestemt tilstand gjennom det entropiske bidraget for de raskeste bevegelsene28, og proteinfunksjonen kan opprettholdes av bevegelser på forskjellige tidsskalaer fra sub-ps for lysfølsomme proteiner29 til ms for domenebevegelser, noe som kan tilrettelegges av picosekund-nanosekunddynamikk30.
To eksempler på bruk av nøytron-tilbakespredningsspektroskopi for å studere amyloidproteiner vil bli presentert, en i flytende tilstand for å studere proteindynamikk og en i hydrert pulvertilstand for å studere hydreringsvanndynamikk. Det første eksemplet gjelder aggregering av lysozym i μm størrelse sfærer (kalt partikler) fulgt i sanntid5, og den andre en sammenligning av vanndynamikk i innfødte og aggregerte tilstander av det humane protein tau31.
Lysozym er et enzym involvert i immunforsvar og består av 129 aminosyrerester. Lysozym kan danne partikler i deuterert buffer ved pD på 10,5 og ved en temperatur på 90 °C. Med nøytronspredning viste vi at tidsutviklingen av lysozymets senter-av-massediffusjonskoeffisient følger den enkle eksponentielle kinetikken til tioflavin T-fluorescens (en fluorescerende sonde som brukes til å overvåke dannelsen av amyloid kryss-β mønstre32), noe som indikerer at dannelsespartikkelformige overbygninger og kryss-β-mønstre forekommer i et enkelt trinn med samme hastighet. Videre forble den interne dynamikken konstant gjennom aggregeringsprosessen, noe som kan forklares enten ved en rask konformasjonsendring som ikke kan observeres på NBS-instrumenter, eller ved fravær av signifikant endring i proteinintern energi ved aggregering.
Det humane proteinet tau er et egenordnet protein (IDP) som består av 441 aminosyrer for den såkalte 2N4R isoformen, som er spesielt involvert i Alzheimers sykdom33. Ved å bruke nøytron-backscattering på pulver av perdeuterert protein tau, viste vi at hydreringsvanndynamikken økes i fibertilstanden, med en høyere populasjon av vannmolekyler som gjennomgår translasjonsbevegelser. Resultatet antyder at en økning i hydreringsvannentropi kan drive amyloidflimmer av tau.
