El neutrón es una partícula masiva y sin carga que se ha utilizado con éxito a lo largo de los años para sondear muestras en diversos campos, desde la física fundamental hasta la biología1. Para aplicaciones biológicas, la dispersión de neutrones de ángulo pequeño, la dispersión inelástica de neutrones y la cristalografía y reflectometría de neutrones se utilizan ampliamente 2,3,4. La dispersión inelástica de neutrones proporciona una medición promediada por conjuntos de la dinámica sin requerir un etiquetado específico per se, y una calidad de señal que no depende del tamaño o la proteína5. La medición se puede hacer utilizando un ambiente altamente complejo para la proteína en estudio que imita el medio intracelular, como un lisado bacteriano deuterado o incluso in vivo 3,6,7. Se pueden usar diferentes configuraciones experimentales para estudiar la dinámica, a saber, i) acceso de tiempo de vuelo a la dinámica sub-ps-ps, ii) acceso de retrodispersión a la dinámica ps-ns, y iii) acceso de spin-echo a la dinámica de ns a cientos de ns. La retrodispersión de neutrones hace uso de la ley de Bragg 2d sinθ = nλ, donde d es la distancia entre planos en un cristal, θ el ángulo de dispersión, n el orden de dispersión y λ la longitud de onda. El uso de cristales para la retrodispersión hacia los detectores permite lograr una alta resolución en energía, típicamente ~ 0.8 μeV. Para medir el intercambio de energía, se utiliza una unidad Doppler que lleva un cristal en retrodispersión para definir y ajustar la longitud de onda de neutrones entrantes 8,9,10 (Figura 1), o se puede usar una configuración de tiempo de vuelo a costa de una disminución en la resolución de energía 11.
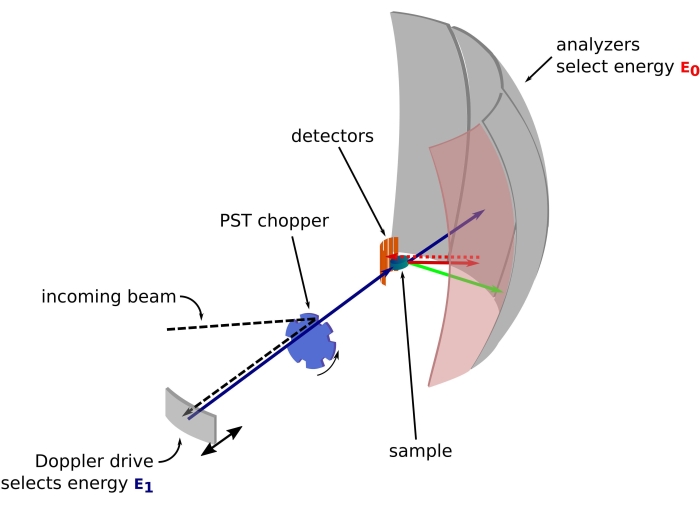
Figura 1: Croquis de un espectrómetro de retrodispersión de neutrones con una unidad Doppler. El haz entrante golpea el helicóptero de transformación del espacio de fase (PST)42, lo que aumenta el flujo en la posición de la muestra. Luego se retrodispersa hacia la muestra mediante la unidad Doppler, que selecciona una energía E1 (flecha cian). Los neutrones son dispersados por la muestra (con diferentes energías representadas por el color de las flechas) y los analizadores, hechos de cristales de Si 111, solo dispersarán neutrones con una energía específica E0 (flechas de color rojo aquí). Por lo tanto, la transferencia de momento q se obtiene de la posición detectada del neutrón en la matriz del detector, y la transferencia de energía se obtiene de la diferencia E1– E0. El tiempo de vuelo esperado para el pulso de neutrones producido por el PST se utiliza para descartar la señal de los neutrones dispersos directamente hacia los tubos detectores. Abreviatura: PST = transformación del espacio de fase. Haga clic aquí para ver una versión más grande de esta figura.
Para la espectroscopia de retrodispersión, la principal contribución a la señal de muestras ricas en protones de hidrógeno, como las proteínas, proviene de la dispersión incoherente, para la cual la intensidad de dispersión Sinc(q, ω) se muestra mediante Eq (1)12
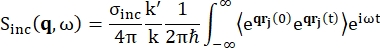 (1)
(1)
Donde σinc es la sección transversal incoherente del elemento considerado, k’ es la norma del vector de onda disperso, k la norma del vector de onda entrante, q (= k – k’) la transferencia de momento, r j(t) el vector de posición del átomo j en el tiempo t, y ω la frecuencia correspondiente a la transferencia de energía entre el neutrón entrante y el sistema. Los corchetes angulares denotan el promedio del conjunto. Por lo tanto, la dispersión incoherente sondea la autocorrelación de partículas individuales promediada por conjuntos de las posiciones de los átomos con el tiempo y da la autodinámica promediada sobre todos los átomos en el sistema y diferentes orígenes de tiempo (promedio de conjunto). La función de dispersión es la transformada de Fourier en el tiempo de la función de dispersión intermedia I(q, t), que puede ser vista como la transformada de Fourier en el espacio de la función de correlación de van Hove mostrada por Eq (2):
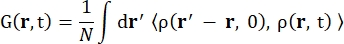 (2)
(2)
Donde ρ(r,t) es la densidad de probabilidad de encontrar un átomo en la posición r y el tiempo t 13.
Para un proceso de difusión fickiano, la función de autodifusión resulta (ver Eq (3)) después de una doble transformada de Fourier en una función de dispersión que consiste en un lorentziano de ancho de línea dado por γ = Dq2.
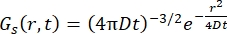 (3)
(3)
Se desarrollaron modelos más sofisticados que se encontraron útiles, como el modelo de difusión de salto de Singwi y Sjölander para la dinámica interna de proteínas ps-ns14 o el modelo de rotación de Sears para el agua de hidratación15,16,17.
En el instrumento de retrodispersión de neutrones (NBS) IN16B8,9 en el ILL, Grenoble, Francia (Figura suplementaria S1), una configuración comúnmente utilizada con proteínas consiste en cristales de Si 111 para los analizadores con un accionamiento Doppler para ajustar la longitud de onda entrante (Figura suplementaria S2A), dando así acceso al rango de transferencia de momento ~0.2 Å-1 < q < ~2 Å-1 y rango de transferencia de energía de –30 μeV < 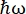 < 30 μeV- correspondiente a escalas de tiempo que van desde unos pocos ps a unos pocos ns y distancias de unos pocos Å. Además, IN16B ofrece la posibilidad de realizar escaneos elásticos e inelásticos de ventana fija (E/IFWS)10, que incluyen la adquisición de datos en una transferencia de energía fija. Como el flujo es limitado cuando se trabaja con neutrones, E/IFWS permite maximizar el flujo para una transferencia de energía, reduciendo así el tiempo de adquisición necesario para obtener una relación señal-ruido satisfactoria. Una opción más reciente es el modo11 del espectrómetro de retrodispersión y tiempo de vuelo (BATS), que permite la medición de una amplia gama de transferencias de energía (por ejemplo, -150 μeV < < 150 μeV), con un flujo más alto que con el accionamiento Doppler, pero a costa de una resolución de energía más baja (Figura suplementaria S2B).
< 30 μeV- correspondiente a escalas de tiempo que van desde unos pocos ps a unos pocos ns y distancias de unos pocos Å. Además, IN16B ofrece la posibilidad de realizar escaneos elásticos e inelásticos de ventana fija (E/IFWS)10, que incluyen la adquisición de datos en una transferencia de energía fija. Como el flujo es limitado cuando se trabaja con neutrones, E/IFWS permite maximizar el flujo para una transferencia de energía, reduciendo así el tiempo de adquisición necesario para obtener una relación señal-ruido satisfactoria. Una opción más reciente es el modo11 del espectrómetro de retrodispersión y tiempo de vuelo (BATS), que permite la medición de una amplia gama de transferencias de energía (por ejemplo, -150 μeV < < 150 μeV), con un flujo más alto que con el accionamiento Doppler, pero a costa de una resolución de energía más baja (Figura suplementaria S2B).
Una propiedad importante de la dispersión de neutrones es que la sección transversal incoherente σinc tiene un valor 40 veces mayor para el hidrógeno que para el deuterio y es insignificante para otros elementos que se encuentran comúnmente en muestras biológicas. Por lo tanto, la dinámica de las proteínas en un ambiente líquido se puede estudiar mediante el uso de un tampón deuterado, y el estado del polvo permite el estudio de la dinámica interna de la proteína con proteína en polvo hidrogenada hidratada con D2O, o el estudio del agua de hidratación para la proteína perdeuterada en polvo hidratada conH2O. En estado líquido, la retrodispersión de neutrones permite típicamente el acceso simultáneo a la autodifusión del centro de masa de las proteínas (difusión de tipo fickiano) y su dinámica interna. Estos últimos son movimientos de columna vertebral y de cadena lateral generalmente descritos por el llamado modelo de difusión de salto u otros 3,18. En los polvos de proteína hidrogenada, la difusión de proteínas está ausente y solo se necesita modelar la dinámica interna. Para el agua de hidratación, las contribuciones de los movimientos de traslación y rotación de las moléculas de agua presentan una dependencia diferente de la transferencia de momento q, lo que permite su distinción en el proceso de análisis de datos17.
Este artículo ilustra el método de retrodispersión de neutrones con el estudio de proteínas que se encontraron capaces de desplegarse, agregarse en una forma canónica que consiste en pilas de β hebras -el llamado patrón de β cruzado19,20– y formar fibras alargadas. Se trata de la llamada agregación amiloide, ampliamente estudiada debido a su papel central en trastornos neurodegenerativos como el Alzheimer o el Parkinson21,22. El estudio de las proteínas amiloides también está motivado por el papel funcional que pueden desempeñar 23,24 o su alto potencial para el desarrollo de nuevos biomateriales25. Los determinantes fisicoquímicos de la agregación amiloide siguen sin estar claros, y no se dispone de una teoría general de la agregación amiloide, a pesar del tremendo progreso durante los últimos años21,26.
La agregación amiloide implica cambios en la estructura y estabilidad de las proteínas con el tiempo, cuyo estudio implica naturalmente dinámica, vinculada a la estabilidad de la conformación de proteínas, la función de las proteínas y el paisaje de energía de las proteínas27. La dinámica está directamente relacionada con la estabilidad de un estado específico a través de la contribución entrópica para los movimientos más rápidos28, y la función de la proteína puede ser sostenida por movimientos en varias escalas de tiempo desde sub-ps para proteínas sensibles a la luz29 hasta ms para movimientos de dominio, que pueden ser facilitados por la dinámica de picosegundos-nanosegundos30.
Se presentarán dos ejemplos de uso de la espectroscopia de retrodispersión de neutrones para estudiar proteínas amiloides, uno en estado líquido para estudiar la dinámica de proteínas y otro en estado de polvo hidratado para estudiar la dinámica del agua de hidratación. El primer ejemplo se refiere a la agregación de lisozima en esferas de tamaño μm (llamadas partículas) seguidas en tiempo real5, y el segundo a una comparación de la dinámica del agua en estados nativos y agregados de la proteína humana tau31.
La lisozima es una enzima implicada en la defensa inmune y está compuesta por 129 residuos de aminoácidos. La lisozima puede formar partículas en tampón deuterado a pD de 10,5 y a una temperatura de 90 °C. Con la dispersión de neutrones, demostramos que la evolución temporal del coeficiente de difusión del centro de masa de la lisozima sigue la cinética exponencial única de la fluorescencia de tioflavina T (una sonda fluorescente utilizada para monitorear la formación de patrones de β cruzada amiloide32), lo que indica que la formación de superestructuras particuladas y patrones de β cruzada ocurren en un solo paso con la misma velocidad. Además, la dinámica interna se mantuvo constante durante todo el proceso de agregación, lo que puede explicarse por un cambio conformacional rápido que no se puede observar en los instrumentos NBS, o por la ausencia de un cambio significativo en la energía interna de las proteínas tras la agregación.
La proteína humana tau es una proteína intrínsecamente desordenada (IDP) que consta de 441 aminoácidos para la llamada isoforma 2N4R, que está notablemente involucrada en la enfermedad de Alzheimer33. Usando retrodispersión de neutrones en polvos de proteína tau perdeuterada, demostramos que la dinámica del agua de hidratación aumenta en el estado de fibra, con una mayor población de moléculas de agua sometidas a movimientos de traslación. El resultado sugiere que un aumento en la entropía del agua de hidratación podría conducir a la fibrilación amiloide de tau.
