L’infiltration lipidique des muscles squelettiques, connue sous le nom de myostéatose, augmente avec l’obésité et le vieillissement. La myostéatose est négativement corrélée à la masse musculaire et à la force et à la sensibilité à l’insuline1. De plus, des études récentes indiquent que le degré de myostéatose pourrait être utilisé comme facteur pronostique pour d’autres affections telles que lesmaladies cardiovasculaires 2, la stéatose hépatique non alcoolique3 ou le cancer4. Les lipides peuvent s’accumuler dans le muscle squelettique entre les fibres musculaires sous forme de lipides extramyocellulaires ou dans les fibres, sous forme de lipides intramyocellulaires (IMCL). Les IMCL sont principalement stockés sous forme de triglycérides dans des gouttelettes lipidiques (LD) qui sont utilisées comme carburant métabolique pendant l’exercice physique 5,6. Cependant, lorsque l’offre de lipides dépasse la demande ou lorsque les mitochondries deviennent dysfonctionnelles, les IMCL seront impliqués dans la résistance musculaire à l’insuline, comme on le voit chez les personnes métaboliquement malsaines, obèses et chez les patients diabétiques de type 27. Curieusement, les athlètes d’endurance ont des niveaux similaires, sinon plus élevés, de MCMC que ceux trouvés chez les patients obèses atteints de diabète sucré de type 2, tout en maintenant une sensibilité élevée à l’insuline. Ce phénomène est décrit comme le « paradoxe de l’athlète »8,9, et s’explique par une évaluation plus nuancée des TA musculaires, liée à leur taille, leur densité, leur localisation, leur dynamique et la composition des espèces lipidiques.
Tout d’abord, la taille de la LD est inversement corrélée à la sensibilité à l’insuline et à la forme physique10,11. En fait, les LD plus petites présentent une surface relativement plus grande pour l’action de la lipase et, par conséquent, ont potentiellement une plus grande capacité à mobiliser les lipides12. Deuxièmement, la densité LD (nombre/surface) joue un rôle controversé dans l’actionde l’insuline 8,10; pourtant, il semble être augmenté chez les athlètes. Troisièmement, la localisation subcellulaire des LD est importante, car les LD situées juste sous la membrane superficielle (sous-sarmateuse ou périphérique) exercent un effet plus délétère sur la sensibilité à l’insuline que les LD centrales 8,9,13. Ces dernières fournissent du carburant aux mitochondries centrales, qui ont une plus grande activité respiratoire et sont plus spécialisées pour répondre à la forte demande d’énergie requise pour la contraction14. En revanche, les TA périphériques fournissent des mitochondries sous-sarmateuses, qui sont impliquées dans les processus liés à la membrane8. Enfin, au-delà des triglycérides, des lipides complexes spécifiques dans le muscle peuvent être plus délétères que d’autres. Par exemple, le diacylglycérol, l’acyl-CoA à longue chaîne et les céramides peuvent s’accumuler dans les muscles lorsque le taux de renouvellement des triglycérides est faible, altérant ainsi la signalisation de l’insuline 9,15. Pour en revenir au « paradoxe de l’athlète », les athlètes d’endurance ont un nombre élevé de TA centrales plus petites avec des taux de renouvellement élevés dans les fibres de type I (oxydatives), tandis que les patients obèses et diabétiques ont des TA périphériques plus importants avec de faibles taux de renouvellement dans les fibres de type II (glycolytiques) 8,15,16. En plus de leur rôle dans le stockage et la libération de l’énergie, les LD via des acides gras dérivés (FA) et une protéine de pelage (périlamine 5) pourraient également fonctionner comme des acteurs critiques impliqués dans la régulation transcriptionnelle de l’oxydation de l’AF et de la biogenèse mitochondriale8. En raison de leurs implications cruciales en physiologie et en physiopathologie, des études approfondies sur la dynamique et les fonctions des TA sont justifiées.
Bien qu’il existe plusieurs techniques pour étudier les IMCL, elles ne sont pas toutes adaptées pour quantifier avec précision la taille, la densité et la distribution des DA DL d’une manière spécifique à la fibre. Par exemple, l’évaluation des NML par spectroscopie par résonance magnétique, tout en étant non invasive, offre un niveau de résolution qui n’est pas suffisant pour étudier la taille et l’emplacement précis des LD dans la fibre, et ce n’est pas spécifique au type de fibre17,18. De même, les techniques biochimiques effectuées sur des homogénats musculaires entiers19 ne peuvent pas évaluer l’emplacement et la taille des lipides. Par conséquent, la méthode la plus adéquate pour analyser la morphologie et l’emplacement de la LD est la microscopie électronique à transmission quantitative13, mais cette technique est coûteuse et prend beaucoup de temps. Par conséquent, l’imagerie par fluorescence confocale sur des préparations avec des colorants tels que Oil Red O (ORO)20,21, monodansylpentane (MDH)22 ou Bodipy 23,24,25, est apparue comme le meilleur outil pour ces études.
Ici, un protocole complet est décrit, y compris l’échantillonnage et le traitement des tissus, la coloration corporelle et l’acquisition et l’analyse d’images confocales pour quantifier la taille, le nombre et la localisation des TA dans les cryosections musculaires de souris. Étant donné que les IMCL ne sont pas répartis uniformément entre les fibres oxydatives et glycolytiques, et que chaque type de fibre régule différemment la dynamique de la LD, l’étude des IMCL doit être spécifique au type de fibre 16,25,26,27. Par conséquent, ce protocole utilise l’immunofluorescence sur des sections en série pour identifier les isoformes de chaîne lourde de myosine (MyHC) exprimées par chaque fibre. Un autre avantage de ce protocole est le traitement simultané d’un muscle glycolytique (extenseur digitorum longus, EDL) et d’un muscle oxydatif (soléaire) placé côte à côte avant la congélation (Figure 1). Ce traitement simultané permet non seulement de gagner du temps, mais aussi d’éviter la variabilité due au traitement séparé des échantillons.
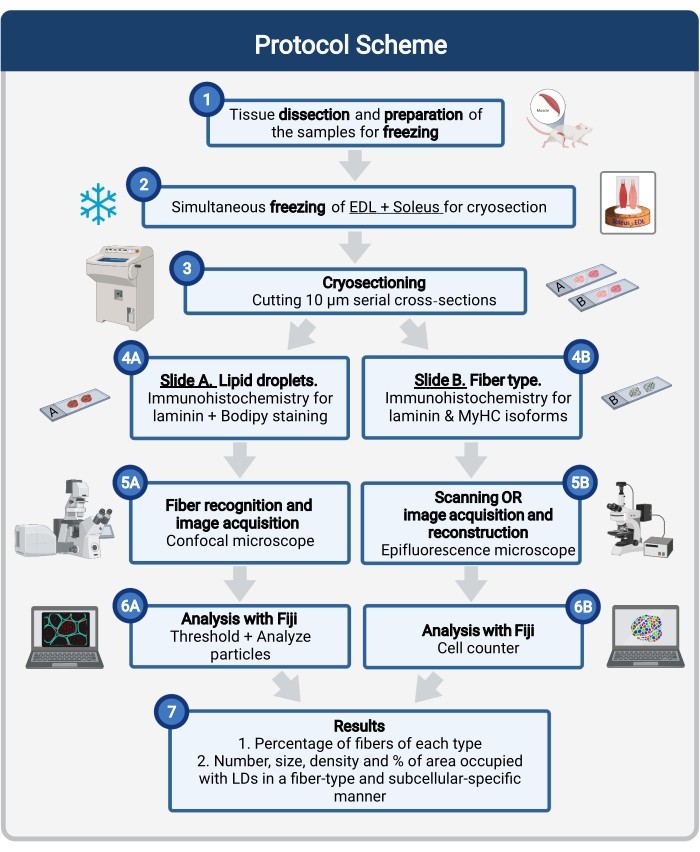
Figure 1 : Vue d’ensemble schématique de la procédure. Après la dissection musculaire (1), des muscles sélectionnés de taille similaire sont préparés et congelés ensemble (2). Les sections transversales en série de 10 μm sont obtenues à l’aide d’un cryostat et directement montées sur des lames d’adhérence (3). À partir de deux diapositives en série, la première (4A) est immunomarquée pour la laminine et colorée avec Bodipy pour reconnaître les LD et la seconde (4B) est immunocolorée avec des anticorps contre les MyHC pour la reconnaissance des types de fibres musculaires. Les images sont acquises à l’aide d’un microscope confocal pour Bodipy (5A) et d’un microscope à épifluorescence pour les types de fibres musculaires (5B). Les images sont analysées aux Fidji en appliquant un seuil et en quantifiant les particules (6A) pour obtenir le nombre, la taille moyenne, la densité et le pourcentage de la surface totale occupée par les TA (7) ou en comptant les cellules (6B) pour obtenir le pourcentage de fibres de chaque type dans la section (7). Abréviations : LD = gouttelettes lipidiques ; EDL = extenseur digitorum longus; MyHC = isoformes de chaîne lourde de myosine. Veuillez cliquer ici pour voir une version agrandie de cette figure.
