A infiltração de lipídios musculares esqueléticos, conhecida como micosteatose, aumenta com obesidade e envelhecimento. A miostetose está negativamente correlacionada com massa muscular e força e com sensibilidade à insulina1. Além disso, estudos recentes indicam que o grau de mostetose poderia ser usado como fator prognóstico para outras condições como doença cardiovascular2, doença hepática gordurosa não alcoólica3 ou câncer4. Lipídios podem se acumular no músculo esquelético entre as fibras musculares como lipídios extramyocelulares ou dentro das fibras, como lipídios intramyocelulares (IMCLs). Os IMCLs são predominantemente armazenados como triglicérides em gotículas lipídicas (LDs) que são usadas como combustível metabólico durante o exercício físico 5,6. No entanto, quando a oferta lipídica excede a demanda, ou quando as mitocôndrias se tornarem disfuncionais, os IMCLs serão implicados na resistência à insulina muscular, como visto em indivíduos metabolicamente insalubres, obesos e em pacientes com diabetes tipo 27. Curiosamente, os atletas de resistência têm níveis semelhantes, se não mais elevados, de IMCLs aos encontrados em pacientes obesos com diabetes mellitus tipo 2, mantendo alta sensibilidade à insulina. Esse fenômeno é descrito como o “paradoxo do atleta”8,9, e é explicado por uma avaliação mais matizada de LDs musculares, relacionadas ao seu tamanho, densidade, localização, dinâmica e composição de espécies lipídicas.
Primeiro, o tamanho do LD está inversamente correlacionado à sensibilidade à insulina e ao condicionamento físico10,11. De fato, LDs menores exibem uma área de superfície relativamente maior para a ação de lipase e, portanto, potencialmente têm maior capacidade de mobilizar lipídios12. Em segundo lugar, a densidade LD (número/superfície) desempenha um papel controverso na ação de insulina 8,10; ainda, parece ser aumentado em atletas. Em terceiro lugar, a localização subcelular de LDs é importante, uma vez que os LDs localizados logo abaixo da membrana superficial (subsarcolemmal ou periférico) exercem um efeito mais deletério sobre a sensibilidade à insulina do que os centrais 8,9,13. Estas últimas fornecem combustível para mitocôndrias centrais, que possuem maior atividade respiratória e são mais especializadas para atender à alta demanda energética necessária para a contração14. Em contrapartida, os LDs periféricos fornecem mitocôndrias subsarcolemmal, que estão envolvidas nos processos relacionados à membrana8. Finalmente, além dos triglicérides, lipídios complexos específicos dentro do músculo podem ser mais deletérios do que outros. Por exemplo, diaciglicerol, acicl-CoA de cadeia longa e ceramidas podem se acumular em músculos quando a taxa de rotatividade de triglicérides é baixa, prejudicando assim a sinalização de insulina 9,15. Voltando ao “paradoxo do atleta”, os atletas de resistência têm um alto número de LDs centrais menores com taxas de rotatividade elevadas nas fibras tipo I (oxidativa), enquanto pacientes obesos e diabéticos têm LDs periféricos maiores com baixas taxas de rotatividade nas fibras tipo II (glicolítico) 8,15,16. Além de seu papel no armazenamento e liberação de energia, os LDs via ácidos graxos derivados (FA) e uma proteína de casaco (perilipin 5) também poderiam funcionar como atores críticos envolvidos na regulação transcricional da oxidação da FA e biogênese mitocondrial8. Devido às suas implicações cruciais na fisiologia e na fisiopatologia, estudos aprofundados sobre dinâmicas e funções de LDs são justificados.
Embora existam várias técnicas para estudar IMCLs, elas não são todas adequadas para quantificar com precisão o tamanho, a densidade e a distribuição de LD de forma específica. Por exemplo, a avaliação de IMCLs por espectroscopia de ressonância magnética, embora não invasiva, oferece um nível de resolução que não é suficiente para estudar o tamanho e a localização precisa de LDs dentro da fibra, e não é específica do tipo fibra 17,18. Da mesma forma, técnicas bioquímicas realizadas em homogeneizadores muscularesinteiros 19 não podem avaliar a localização e o tamanho dos lipídios. Consequentemente, o método mais adequado para analisar a morfologia e localização de LD é a microscopia eletrônica de transmissão quantitativa13, mas essa técnica é cara e demorada. Portanto, a fluorescência confocal em preparações com corantes como Oil Red O (ORO)20,21, monodansylpentane (MDH)22, ou Bodipy 23,24,25, emergiu como a melhor ferramenta para esses estudos.
Aqui, é descrito um protocolo completo, incluindo amostragem e processamento de tecidos, coloração bodipy e aquisição e análise de imagem confocal para quantificar o tamanho, o número e a localização de LD em crioseções musculares de camundongos. Uma vez que as IMCLs não são distribuídas uniformemente entre fibras oxidativas e glicolíticas, e cada tipo de fibra regula a dinâmica LD de forma diferente, o estudo das IMCLs deve ser específico do tipo fibra 16,25,26,27. Portanto, este protocolo utiliza imunofluorescência em seções seriais para identificar isoform(s) de cadeia pesada de miosina (MyHC) expressa por cada fibra. Outra vantagem deste protocolo é o processamento simultâneo de um músculo glicolítico (extensor digitorum longus, EDL) e um músculo oxidativo (soleus) colocado lado a lado antes de congelar (Figura 1). Esse processamento simultâneo não só economiza tempo, mas também evita a variabilidade devido ao processamento separado das amostras.
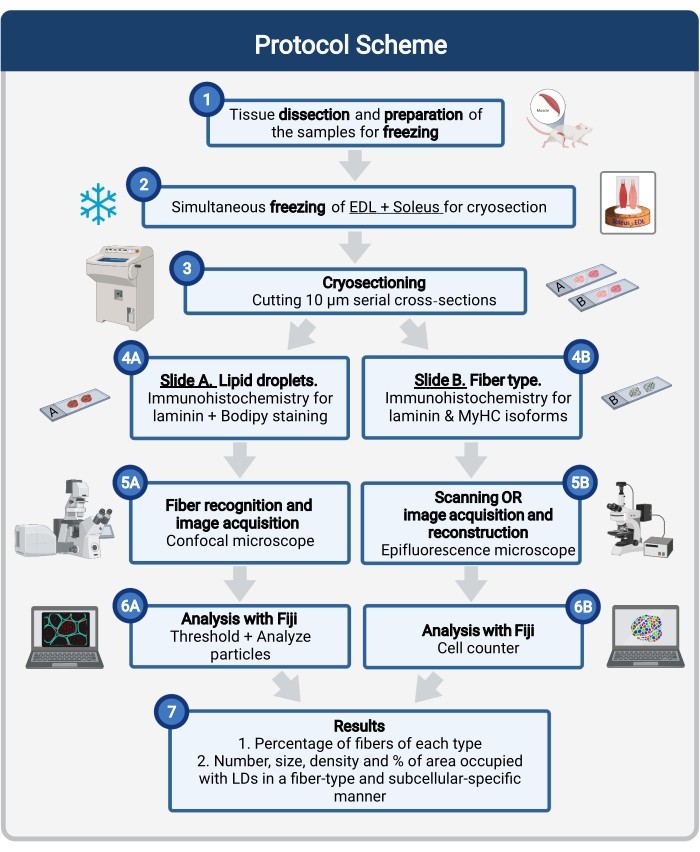
Figura 1: Visão geral do procedimento. Após dissecção muscular (1), músculos selecionados de tamanho semelhante são preparados e congelados juntos (2). Seções transversais em série de 10 μm são obtidas usando um criostat e montadas diretamente em slides de adesão (3). A partir de dois slides seriais, o primeiro (4A) é imunolabeled para laminina e manchado com Bodipy para reconhecer LDs e o segundo (4B) é imunossumado com anticorpos contra MyHCs para o reconhecimento de tipos de fibras musculares. As imagens são adquiridas usando um microscópio confocal para Bodipy (5A) e um microscópio de epifluorescência para tipos de fibras musculares (5B). As imagens são analisadas em Fiji aplicando um limiar e quantificando partículas (6A) para obter o número, tamanho médio, densidade e percentual da área total ocupada por LDs (7) ou células de contagem (6B) para obter a porcentagem de fibras de cada tipo na seção (7). Abreviaturas: LDs = gotículas lipídicas; EDL = extensor digitorum longus; MyHCs = isoforms de cadeia pesada de miosina. Clique aqui para ver uma versão maior desta figura.
