La infiltración de lípidos del músculo esquelético, conocida como miosteatosis, aumenta con la obesidad y el envejecimiento. La miostetosis se correlaciona negativamente con la masa muscular y la fuerza y con la sensibilidad a la insulina1. Además, estudios recientes indican que el grado de miosteatosis podría usarse como factor pronóstico para otras afecciones como la enfermedad cardiovascular2, la enfermedad del hígado graso no alcohólico3 o el cáncer4. Los lípidos pueden acumularse en el músculo esquelético entre las fibras musculares como lípidos extramiocelulares o dentro de las fibras, como lípidos intramiocelulares (MMCL). Los MMCL se almacenan predominantemente como triglicéridos en gotas lipídicas (LD) que se utilizan como combustible metabólico durante el ejercicio físico 5,6. Sin embargo, cuando el suministro de lípidos excede la demanda, o cuando las mitocondrias se vuelven disfuncionales, las MMCL estarán implicadas en la resistencia muscular a la insulina, como se ve en individuos obesos metabólicamente insalubres y en pacientes con diabetes tipo 27. Curiosamente, los atletas de resistencia tienen niveles similares, si no más altos, de MMCL a los que se encuentran en pacientes obesos con diabetes mellitus tipo 2, mientras que mantienen una alta sensibilidad a la insulina. Este fenómeno se describe como la “paradoja del atleta”8,9, y se explica por una evaluación más matizada de los LD musculares, relacionados con su tamaño, densidad, localización, dinámica y composición de especies lipídicas.
En primer lugar, el tamaño de la DA está inversamente correlacionado con la sensibilidad a la insulina y la aptitud física10,11. De hecho, los LD más pequeños exhiben un área de superficie relativamente mayor para la acción de la lipasa y, por lo tanto, potencialmente tienen una mayor capacidad para movilizar lípidos12. En segundo lugar, la densidad de LD (número/superficie) juega un papel controvertido en la acción de la insulina 8,10; sin embargo, parece estar aumentado en los atletas. En tercer lugar, la localización subcelular de los LD es importante, ya que los LD situados justo debajo de la membrana superficial (subsarcolémmica o periférica) ejercen un efecto más perjudicial sobre la sensibilidad a la insulina que los centrales 8,9,13. Estos últimos proporcionan combustible a las mitocondrias centrales, que tienen una mayor actividad respiratoria y están más especializadas para satisfacer la alta demanda de energía requerida para la contracción14. Por el contrario, los LD periféricos suministran mitocondrias subsarcolemmales, que están involucradas en procesos relacionados con la membrana8. Finalmente, más allá de los triglicéridos, los lípidos complejos específicos dentro del músculo pueden ser más perjudiciales que otros. Por ejemplo, el diacilglicerol, el acil-CoA de cadena larga y las ceramidas pueden acumularse en el músculo cuando la tasa de renovación de triglicéridos es baja, lo que afecta la señalización de la insulina 9,15. Volviendo a la “paradoja del atleta”, los atletas de resistencia tienen un alto número de LD centrales más pequeños con tasas de rotación elevadas en fibras tipo I (oxidativas), mientras que los pacientes obesos y diabéticos tienen LD periféricos más grandes con bajas tasas de renovación en fibras tipo II (glucolíticas) 8,15,16. Además de su papel en el almacenamiento y liberación de energía, los LD a través de ácidos grasos derivados (FA) y una proteína de capa (perilipina 5) también podrían funcionar como actores críticos involucrados en la regulación transcripcional de la oxidación de FA y la biogénesis mitocondrial8. Debido a sus implicaciones cruciales en fisiología y fisiopatología, se justifican estudios en profundidad sobre la dinámica y las funciones de los LD.
Aunque existen varias técnicas para estudiar las IGMCL, no todas son adecuadas para cuantificar con precisión el tamaño, la densidad y la distribución de la LD de una manera específica de la fibra. Por ejemplo, la evaluación de los MMCL por espectroscopia de resonancia magnética, si bien no es invasiva, ofrece un nivel de resolución que no es suficiente para estudiar el tamaño y la ubicación precisa de los LD dentro de la fibra, y no es específico del tipo de fibra17,18. Del mismo modo, las técnicas bioquímicas realizadas en homogeneizados de músculo entero19 no pueden evaluar la ubicación y el tamaño de los lípidos. En consecuencia, el método más adecuado para analizar la morfología y la ubicación de la DA es la microscopía electrónica de transmisión cuantitativa13, pero esta técnica es costosa y requiere mucho tiempo. Por lo tanto, la imagen de fluorescencia confocal en preparaciones con colorantes como Oil Red O (ORO)20,21, monodansilipentano (MDH)22 o Bodipy 23,24,25, se ha convertido en la mejor herramienta para estos estudios.
Aquí, se describe un protocolo completo, que incluye muestreo y procesamiento de tejidos, tinción de Bodipy y adquisición y análisis de imágenes confocales para cuantificar el tamaño, el número y la localización de LD en criosecciones musculares de ratón. Dado que los MMCL no están distribuidos uniformemente entre las fibras oxidativas y glucolíticas, y cada tipo de fibra regula la dinámica de la LD de manera diferente, el estudio de los MMCL debe ser específico del tipo de fibra 16,25,26,27. Por lo tanto, este protocolo utiliza inmunofluorescencia en secciones seriadas para identificar las isoformas de cadena pesada de miosina (MyHC) expresadas por cada fibra. Otra ventaja de este protocolo es el procesamiento simultáneo de un músculo glicolítico (extensor digitorum longus, EDL) y oxidativo (sóleo) colocados uno al lado del otro antes de la congelación (Figura 1). Este procesamiento simultáneo no solo ahorra tiempo, sino que también evita la variabilidad debido al procesamiento separado de las muestras.
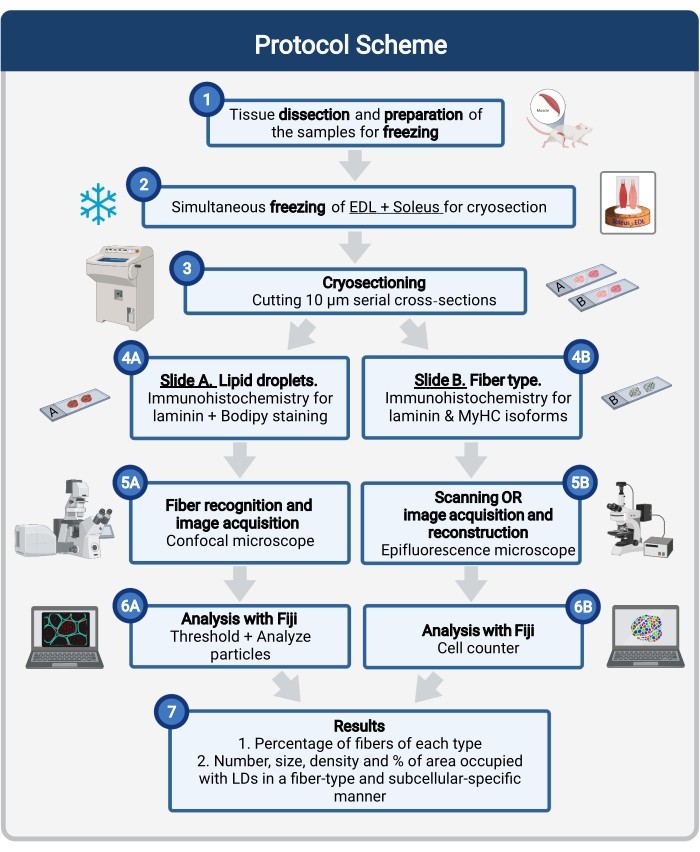
Figura 1: Descripción general esquemática del procedimiento. Después de la disección muscular (1), los músculos seleccionados de tamaño similar se preparan y congelan juntos (2). Las secciones transversales en serie de 10 μm se obtienen mediante un criostato y se montan directamente en portaobjetos de adhesión (3). A partir de dos diapositivas seriadas, la primera (4A) está inmunomarcada para laminina y teñida con Bodipy para reconocer las LD y la segunda (4B) está inmunoteñida con anticuerpos contra MyHCs para el reconocimiento de tipos de fibras musculares. Las imágenes se adquieren utilizando un microscopio confocal para Bodipy (5A) y un microscopio de epifluorescencia para tipos de fibras musculares (5B). Las imágenes se analizan en Fiji aplicando un umbral y cuantificando partículas (6A) para obtener el número, tamaño medio, densidad y porcentaje del área total ocupada por LD (7) o celdas de conteo (6B) para obtener el porcentaje de fibras de cada tipo en la sección (7). Abreviaturas: LD = gotas lipídicas; EDL = extensor digitorum longus; MyHCs = isoformas de cadena pesada de miosina. Haga clic aquí para ver una versión más grande de esta figura.
