タンパク質リン酸化は、DNA損傷に対する応答、成長因子シグナル伝達、および有糸分裂1,2,3の通過を含むがこれらに限定されない、ほとんどの細胞プロセスを制御する。哺乳動物細胞では、タンパク質の大部分は、ある時点で1つ以上のセリン、スレオニン、またはチロシン残基でリン酸化され、ホスホセリンおよびホスホスレオニンは、すべてのリン酸化部位の約98%を占める2,3。キナーゼは細胞シグナル伝達において広く研究されてきたが、動的細胞プロセスの調節におけるPPPの役割はまだ現れている。
リン酸化ダイナミクスは、キナーゼとホスファターゼの間の動的相互作用によって制御される。哺乳類細胞には、セリン/スレオニンリン酸化を触媒する400以上のプロテインキナーゼが存在する。これらの部位の90%以上は、PP1、PP2A、PP2B、PP4-7、PPT、およびPPZ 2,3からなる酵素の小さなファミリーであるリンタンパク質ホスファターゼ(PPP)によって脱リン酸化されている。PP1およびPP2Aは、細胞内のホスホセリンおよびホスホスレオニン脱リン酸化の大部分を担う2、3、4。キナーゼとホスファターゼの数の顕著な違いおよびインビトロでのPPP触媒サブユニットの特異性の欠如は、キナーゼがリン酸化の主な決定因子であるという信念につながった2,3。しかしながら、複数の研究は、ホスファターゼが多量体ホロ酵素5、6、7、8、9の形成を通じて基質特異性を確立することを示している。例えば、PP1は、触媒サブユニットおよび、所与の時点で、150を超える調節サブユニットのうちの1つからなるヘテロ二量体である6、7、8である。逆に、PP2Aは、足場(A)、調節(B)、および触媒(C)サブユニット2,3,9から形成されるヘテロ三量体である。PP2A調節サブユニット(B55、B56、PR72、および線条体)の4つの異なるファミリーがあり、それぞれに複数の遺伝子、スプライスバリアント、および局在化パターン2、3、9がある。PPPの多量体性は、キナーゼおよびPPP触媒サブユニットの数のギャップを埋める。しかし、PPPシグナリングを研究するための分析上の課題が生じます。PPPシグナル伝達を網羅的に解析するためには、細胞または組織内の様々なホロ酵素を調べることが重要です。キナーゼ阻害剤ビーズ(マルチプレックス阻害剤ビーズまたはキノビーズと呼ばれる)の使用を通じてヒトキノメの研究において大きな進歩がなされており、キナーゼ阻害剤がビーズに固定化され、質量分析法が濃縮キナーゼおよびそれらのインターアクターを同定するために使用される化学的プロテオミクス戦略10,11,12,13。
我々は、PPP生物学を研究するために同様のアプローチを確立している。この技術は、ホスファターゼ阻害剤ビーズ(PIB)と呼ばれるミクロシスチン−LR(MCLR)と呼ばれる固定化された非選択的PPP阻害剤を有するビーズを用いたPPP触媒サブユニットの親和性捕捉を含む14,15。タンパク質活性または局在化を変化させる可能性のある外因性PPPサブユニットの内因性タグ付けまたは発現を必要とする他の方法とは異なり、PIB-MSは、内因性PPP触媒サブユニット、それらに関連する調節および足場サブユニット、および所定の時点または特定の治療条件下での細胞および組織からの相互作用タンパク質(PPPomeと呼ばれる)の濃縮を可能にする。MCLRは、PP1、PP2A、PP4-6、PPT、およびPPZをナノモル濃度で阻害し、PIBをPPPome16の濃縮に非常に効果的にする。この方法は、細胞から臨床サンプルまでの任意の出発物質に使用するためにスケーリングすることができる。ここでは、内因性PPPomeとその修飾状態を効率的に捕捉、同定、定量化するためのPIBおよび質量分析(PIB-MS)の使用について詳しく説明します。
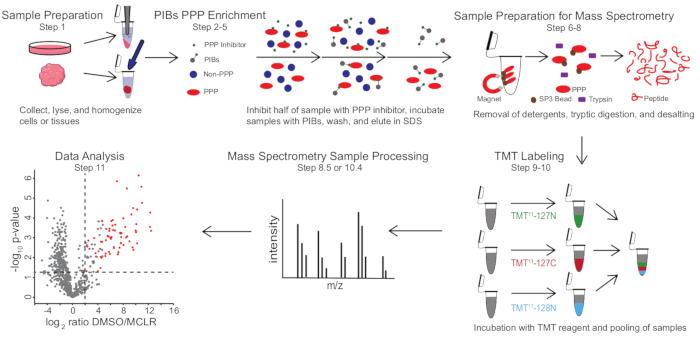
図1:PIB-MSプロトコルの視覚的な概要 PIB-MS実験では、細胞から腫瘍まで、さまざまな形態でサンプルを得ることができます。サンプルは、PPP 濃縮の前に収集、溶解、および均質化されます。PPPを富化するために、溶解液をMCLRなどのPPP阻害剤の有無にかかわらずPIBと共にインキュベートする。その後、PIBを洗浄し、PPPを変性条件下で溶出します。サンプルは、SP3タンパク質濃縮、トリプシン消化、および脱塩による洗剤の除去によって質量分析分析用に調製される。次いで、サンプルを、質量分析分析の前に任意選択でTMT標識することができる。 この図の拡大版を表示するには、ここをクリックしてください。
PIB-MSには、細胞または組織の溶解および清澄化、溶解液とPIBとのインキュベーション、溶出、およびウェスタンブロッティングまたは質量分析ベースのアプローチによる溶出液の分析が含まれます(図1)。遊離MCLRの添加は、特異的PIB結合剤を非特異的相互作用体から区別するための対照として使用することができる。ほとんどのアプリケーションでは、溶出液中のタンパク質を直接同定するためにラベルフリーのアプローチを使用できます。定量化または低存在種の同定においてより高い精度が必要な場合は、タンデム質量タグ(TMT)標識によるさらなる処理を使用して、カバレッジを増加させ、入力を減少させることができる。
