تتحكم فسفرة البروتين في معظم العمليات الخلوية ، بما في ذلك على سبيل المثال لا الحصر الاستجابة لتلف الحمض النووي ، وإشارات عامل النمو ، والمرور عبر الانقسام1،2،3. في خلايا الثدييات ، يتم فسفوريلات غالبية البروتينات في واحد أو أكثر من بقايا سيرين أو ثريونين أو التيروزين في وقت ما ، مع فوسفوسرين وفوسفوثريونين يشكلون حوالي 98 ٪ من جميع مواقع الفسفرة 2,3. في حين تمت دراسة الكينازات على نطاق واسع في الإشارات الخلوية ، فإن دور الشراكات بين القطاعين العام والخاص في تنظيم العمليات الخلوية الديناميكية لا يزال ناشئا.
يتم التحكم في ديناميكيات الفسفرة من خلال التفاعل الديناميكي بين الكينازات والفوسفاتيز. في خلايا الثدييات ، هناك أكثر من 400 كيناز بروتين يحفز فسفرة سيرين / ثريونين. أكثر من 90٪ من هذه المواقع يتم نزع الفوسفوريلات بواسطة فوسفاتيز البروتين الفوسفوبروتيني (PPPs) ، وهي عائلة صغيرة من الإنزيمات التي تتكون من PP1 و PP2A و PP2B و PP4-7 و PPT و PPZ 2,3. PP1 و PP2A مسؤولان عن غالبية إزالة الفوسفوسرين والفوسفوثريونين داخل الخلية2،3،4. أدى الاختلاف الملحوظ في العدد بين الكينازات والفوسفاتيز وعدم خصوصية الوحدات الفرعية الحفازة PPP في المختبر إلى الاعتقاد بأن الكينازات هي المحدد الرئيسي للفسفرة 2,3. ومع ذلك ، فقد أظهرت دراسات متعددة أن الفوسفاتيز يؤسس خصوصية الركيزة من خلال تكوين إنزيمات هولو متعددة الميريك5،6،7،8،9. على سبيل المثال ، PP1 هو غير متجانس يتكون من وحدة فرعية حفازة ، وفي وقت معين ، واحد من أكثر من 150 وحدة فرعية تنظيمية 6,7,8. على العكس من ذلك ، PP2A هو متغاير يتكون من سقالات (A) ، ووحدة تنظيمية (B) ، ووحدة فرعية حفازة (C) 2,3,9. هناك أربع عائلات متميزة من الوحدات الفرعية التنظيمية PP2A (B55 و B56 و PR72 و striatin) ، لكل منها جينات متعددة ، ومتغيرات الربط ، وأنماط التوطين2،3،9. وتسد الطبيعة المتعددة للشراكات بين القطاعين العام والخاص الفجوة في عدد الكينازات والوحدات الفرعية الحفازة لتعادل القوة الشرائية. ومع ذلك ، فإنه يخلق تحديات تحليلية لدراسة إشارات الشراكة بين القطاعين العام والخاص. لتحليل إشارات PPP بشكل شامل ، من الأهمية بمكان التحقيق في مختلف الإنزيمات الهولونية داخل الخلية أو الأنسجة. تم إحراز تقدم كبير في دراسة الكينومي البشري من خلال استخدام حبات مثبطات الكيناز ، التي يطلق عليها اسم حبات مثبطات الإرسال المتعددة أو الخرز ، وهي استراتيجية بروتينية كيميائية حيث يتم تجميد مثبطات الكيناز على الخرز ويستخدم قياس الطيف الكتلي لتحديد الكينازات المخصبة وتفاعلها10،11،12،13.
لقد أنشأنا نهجا مشابها لدراسة بيولوجيا الشراكة بين القطاعين العام والخاص. تتضمن هذه التقنية التقاط التقارب للوحدات الفرعية الحفازة PPP باستخدام الخرز مع مثبط PPP غير انتقائي غير متحرك يسمى microcystin-LR (MCLR) يسمى حبات مثبطات الفوسفاتيز (PIBs) 14,15. على عكس الطرق الأخرى التي تتطلب وضع علامات داخلية أو التعبير عن الوحدات الفرعية لتعادل القوة الشرائية الخارجية التي يمكن أن تغير نشاط البروتين أو توطينه ، يسمح PIB-MS بإثراء الوحدات الفرعية المحفزة لتعادل القوة الشرائية الذاتية المنشأ ، والوحدات الفرعية التنظيمية والسقالات المرتبطة بها ، والبروتينات المتفاعلة (تسمى PPPome) من الخلايا والأنسجة في نقطة زمنية معينة أو في ظل ظروف علاج محددة. يمنع MCLR PP1 و PP2A و PP4-6 و PPT و PPZ بتركيزات نانومولية ، مما يجعل PIBs فعالة للغاية في إثراء PPPome16. يمكن تحجيم هذه الطريقة لاستخدامها على أي مادة أولية من الخلايا إلى العينات السريرية. هنا ، نصف بالتفصيل استخدام PIBs وقياس الطيف الكتلي (PIB-MS) لالتقاط وتحديد وقياس PPPome الداخلي وحالات تعديله بكفاءة.
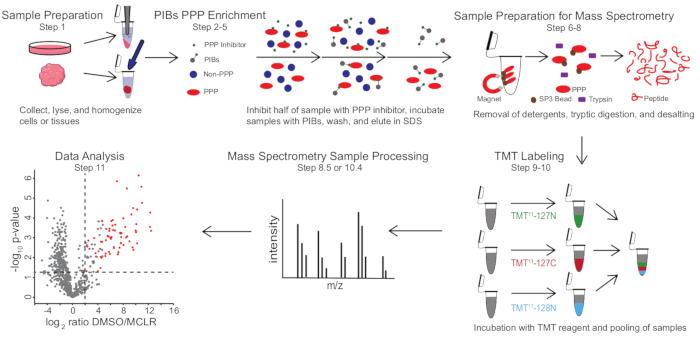
الشكل 1: ملخص مرئي لبروتوكول PIB-MS. في تجربة PIB-MS ، يمكن الحصول على عينات بأشكال مختلفة ، من الخلايا إلى الأورام. يتم جمع العينة وتحليلها وتجانسها قبل إثراء تعادل القوة الشرائية. للإثراء من أجل الشراكات بين القطاعين العام والخاص، يتم احتضان الليزات مع PIBs مع أو بدون مثبط PPP، مثل MCLR. ثم يتم غسل PIBs ، ويتم التخلص من PPPs في ظروف تمسخ. يتم إعداد العينات لتحليل الطيف الكتلي عن طريق إزالة المنظفات من خلال إثراء البروتين SP3 ، والهضم التربتيكي ، وإزالة الملح. يمكن بعد ذلك وضع علامة TMT اختياريا على العينات قبل تحليل مطياف الكتلة. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
يتضمن PIB-MS تحلل وتوضيح الخلايا أو الأنسجة ، وحضانة الليزات باستخدام PIBs ، والاستخلاص ، وتحليل اللوات عبر النشاف الغربي أو النهج القائمة على قياس الطيف الكتلي (الشكل 1). يمكن استخدام إضافة MCLR المجاني كعنصر تحكم لتمييز روابط PIB المحددة عن الجهات الفاعلة غير المحددة. بالنسبة لمعظم التطبيقات ، يمكن استخدام نهج خال من الملصقات لتحديد البروتينات مباشرة في المراوغة. وفي الحالات التي تكون فيها هناك حاجة إلى مزيد من الدقة في التحديد الكمي أو تحديد الأنواع منخفضة الوفرة، يمكن استخدام مزيد من المعالجة باستخدام وسم العلامات الجماعية الترادفية (TMT) لزيادة التغطية وتقليل المدخلات.
