זרחון חלבונים שולט ברוב התהליכים התאיים, כולל אך לא מוגבל לתגובה לנזק לדנ”א, איתות גורמי גדילה ומעבר דרך מיטוזה 1,2,3. בתאי יונקים, רוב החלבונים עוברים זרחון בשאריות סרין, תראונין או טירוזין אחד או יותר בנקודת זמן כלשהי, כאשר פוספוסרינה ופוספוטריאונינים מהווים כ-98% מכלל אתרי הזרחון 2,3. בעוד קינאזות נחקרו בהרחבה באיתות תאי, תפקידם של PPPs בוויסות תהליכים תאיים דינמיים עדיין מתפתח.
דינמיקת הזרחון נשלטת על ידי יחסי הגומלין הדינמיים בין קינאזות לפוספטזות. בתאי יונקים ישנם יותר מ-400 קינאזות חלבוניות המזרזות פוספורילציה של סרין/תראונין. יותר מ-90% מהאתרים האלה עוברים דה-פוספורילציה על-ידי פוספטאזות של פוספופרוטאין (PPPs), משפחה קטנה של אנזימים המורכבת מ-PP1, PP2A, PP2B, PP4-7, PPT ו-PPZ 2,3. PP1 ו-PP2A אחראים לרוב הפוספוסרין והפוספוטראונין דה-פוספורילציה בתוך תא 2,3,4. ההבדל הבולט במספר בין קינאזות לפוספטזות וחוסר הספציפיות של תת-יחידות קטליטיות PPP במבחנה הובילו לאמונה כי קינאזות הן הגורם העיקרי לזרחון 2,3. עם זאת, מחקרים רבים הראו כי פוספטזות מבססות את הספציפיות של המצע באמצעות היווצרות של הולואנזים רב-שמריים 5,6,7,8,9. לדוגמה, PP1 הוא הטרודימר המורכב מיחידת משנה קטליטית, ובזמן נתון, אחד מתוך יותר מ -150 תת-יחידות רגולטוריות 6,7,8. לעומת זאת, PP2A הוא הטרוטרימר שנוצר מפיגומים (A), יחידת משנה רגולטורית (B) ותת-יחידהקטליטית (C) 2,3,9. ישנן ארבע משפחות נפרדות של תת-יחידות רגולטוריות של PP2A (B55, B56, PR72 וסטריאטין), שלכל אחת מהן גנים מרובים, גרסאות שחבור ודפוסי לוקליזציה 2,3,9. האופי הרב-מהותי של PPPs ממלא את הפער במספר קינאזות ותת-יחידות קטליטיות של PPP. עם זאת, היא יוצרת אתגרים אנליטיים לחקר איתות PPP. כדי לנתח באופן מקיף איתות PPP, חשוב לחקור את ההולו-אנזים השונים בתוך תא או רקמה. התקדמות גדולה נעשתה בחקר הקינום האנושי באמצעות שימוש בחרוזי מעכבי קינאז, המכונים חרוזי מעכבי מולטיפלקס או קינוביאדות, אסטרטגיה פרוטאומית כימית שבה מעכבי קינאז משותקים על חרוזים וספקטרומטריית מסה משמשת לזיהוי קינאזות מועשרות והאינטראקצים שלהם 10,11,12,13.
ביססנו גישה דומה לחקר הביולוגיה של PPP. טכניקה זו כוללת לכידת זיקה של תת-יחידות קטליטיות PPP באמצעות חרוזים עם מעכב PPP משותק ולא סלקטיבי הנקרא מיקרוציסטין-LR (MCLR) המכונה חרוזי מעכבי פוספטאז (PIBs)14,15. בניגוד לשיטות אחרות הדורשות תיוג אנדוגני או ביטוי של תת-יחידות PPP אקסוגניות שיכולות לשנות את פעילות החלבון או לוקליזציה שלו, PIB-MS מאפשר העשרה של תת-יחידות קטליטיות אנדוגניות של PPP, תת-יחידות רגולטוריות ופיגומים הקשורות אליהן, וחלבונים מתקשרים (המכונים PPPome) מתאים ורקמות בנקודת זמן נתונה או בתנאי טיפול ספציפיים. MCLR מעכב PP1, PP2A, PP4-6, PPT ו-PPZ בריכוזי ננו-מולארים, מה שהופך את ה-PIBs ליעילים ביותר בהעשרת PPPome16. ניתן לשנות את קנה המידה של שיטה זו לשימוש בכל חומר מוצא, מתאים ועד דגימות קליניות. כאן נתאר בפירוט את השימוש ב-PIBs ובספקטרומטריית מסות (PIB-MS) כדי ללכוד, לזהות ולכמת ביעילות את ה-PPPome האנדוגני ואת מצבי השינוי שלו.
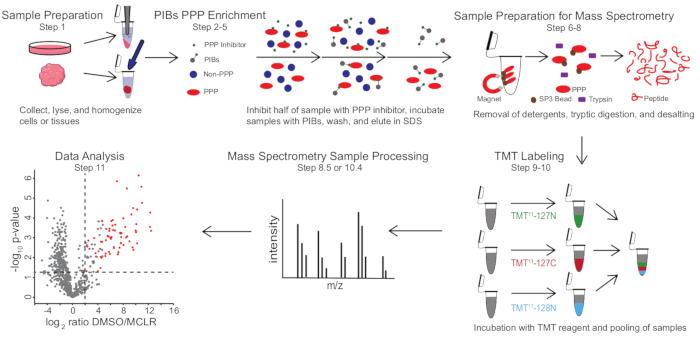
איור 1: סיכום חזותי של פרוטוקול PIB-MS. בניסוי PIB-MS ניתן לקבל דגימות בצורות שונות, מתאים ועד גידולים. הדגימה נאספת, משוקרת ומוגזמת לפני העשרת PPP. כדי להעשיר עבור PPPs, lysate הוא דגירה עם PIBs עם או בלי מעכב PPP, כגון MCLR. לאחר מכן נשטפים ה-PIBs, ו-PPPs נשטפים בתנאי דנטורציה. הדגימות מוכנות לניתוח ספקטרומטריית מסה על ידי הסרת חומרי ניקוי באמצעות העשרת חלבוני SP3, עיכול טריפטי והתפלה. לאחר מכן ניתן לתייג דגימות באופן אופציונלי TMT לפני ניתוח ספקטרומטריית מסות. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
PIB-MS כולל ליזה והבהרה של תאים או רקמות, דגירה של הליזאט עם PIBs, אלוטיון וניתוח של האלואט באמצעות כתמים מערביים או גישות מבוססות ספקטרומטריית מסה (איור 1). התוספת של MCLR חופשי יכולה לשמש כבקרה כדי להבחין בין קלסרים ספציפיים של PIB לבין אינטראקציות לא ספציפיות. עבור רוב היישומים, ניתן להשתמש בגישה נטולת תוויות כדי לזהות ישירות חלבונים ב- eluates. במקרים שבהם יש צורך בדיוק רב יותר בכימות או בזיהוי של מינים בעלי שפע נמוך, ניתן להשתמש בעיבוד נוסף עם תיוג תג מסה טנדם (TMT) כדי להגדיל את הכיסוי ולהפחית את הקלט.
