La fosforilazione proteica controlla la maggior parte dei processi cellulari, tra cui, ma non solo, la risposta al danno al DNA, la segnalazione del fattore di crescita e il passaggio attraverso la mitosi 1,2,3. Nelle cellule di mammifero, la maggior parte delle proteine sono fosforilate in uno o più residui di serina, treonina o tirosina ad un certo punto nel tempo, con fosfosine e fosfotreonine che costituiscono circa il 98% di tutti i siti di fosforilazione 2,3. Mentre le chinasi sono state ampiamente studiate nella segnalazione cellulare, il ruolo dei PPP nella regolazione dei processi cellulari dinamici sta ancora emergendo.
Le dinamiche di fosforilazione sono controllate dall’interazione dinamica tra chinasi e fosfatasi. Nelle cellule di mammifero, ci sono più di 400 protein chinasi che catalizzano la fosforilazione di serina / treonina. Oltre il 90% di questi siti sono defosforilati dalle fosfatasi fosfoproteiche (PPP), una piccola famiglia di enzimi costituita da PP1, PP2A, PP2B, PP4-7, PPT e PPZ 2,3. PP1 e PP2A sono responsabili della maggior parte della fosfoserina e della fosfotreonina defosforilazione all’interno di una cellula 2,3,4. La notevole differenza di numero tra chinasi e fosfatasi e la mancanza di specificità delle subunità catalitiche PPP in vitro hanno portato alla convinzione che le chinasi siano il principale determinante della fosforilazione 2,3. Tuttavia, diversi studi hanno dimostrato che le fosfatasi stabiliscono la specificità del substrato attraverso la formazione di oloenzimi multimerici 5,6,7,8,9. Ad esempio, PP1 è un eterodimero costituito da una subunità catalitica e, in un dato momento, una delle oltre 150 subunità regolatorie 6,7,8. Al contrario, PP2A è un eterotrimero formato da un’impalcatura (A), una subunità regolatoria (B) e una catalitica (C) 2,3,9. Esistono quattro famiglie distinte di subunità regolatorie PP2A (B55, B56, PR72 e striatina), ognuna con più geni, varianti di giunzione e modelli di localizzazione 2,3,9. La natura multimerica dei PPP colma il divario nel numero di chinasi e subunità catalitiche PPP. Tuttavia, crea sfide analitiche per lo studio della segnalazione PPP. Per analizzare in modo completo la segnalazione PPP, è fondamentale studiare i vari oloenzimi all’interno di una cellula o di un tessuto. Grandi progressi sono stati fatti nello studio del kinome umano attraverso l’uso di perline inibitrici della chinasi, chiamate perle o kinobeadi inibitori multiplex, una strategia proteomica chimica in cui gli inibitori della chinasi sono immobilizzati su perline e la spettrometria di massa viene utilizzata per identificare chinasi arricchite e i loro interattori 10,11,12,13.
Abbiamo stabilito un approccio simile per studiare la biologia PPP. Questa tecnica prevede la cattura di affinità di subunità catalitiche PPP utilizzando perline con un inibitore PPP immobilizzato e non selettivo chiamato microcistin-LR (MCLR) chiamato sfere inibitrici della fosfatasi (PIB)14,15. A differenza di altri metodi che richiedono la marcatura endogena o l’espressione di subunità PPP esogene che potrebbero alterare l’attività o la localizzazione delle proteine, PIB-MS consente l’arricchimento di subunità catalitiche PPP endogene, le loro subunità regolatorie e di impalcatura associate e le proteine interagenti (chiamate PPPome) da cellule e tessuti in un dato momento o in condizioni di trattamento specifiche. MCLR inibisce PP1, PP2A, PP4-6, PPT e PPZ a concentrazioni nanomolari, rendendo i PIB altamente efficaci nell’arricchimento per il PPPome16. Questo metodo può essere scalato per l’uso su qualsiasi materiale di partenza dalle cellule ai campioni clinici. Qui, descriviamo in dettaglio l’uso di PIB e spettrometria di massa (PIB-MS) per catturare, identificare e quantificare in modo efficiente il PPPome endogeno e i suoi stati di modifica.
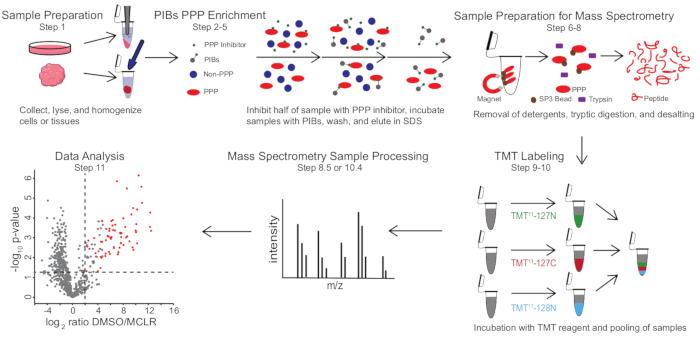
Figura 1: Riepilogo visivo del protocollo PIB-MS. In un esperimento PIB-MS, i campioni possono essere ottenuti in varie forme, dalle cellule ai tumori. Il campione viene raccolto, lisato e omogeneizzato prima dell’arricchimento PPP. Per arricchire i PPP, il lisato viene incubato con PIB con o senza un inibitore PPP, come MCLR. I PIB vengono quindi lavati e i PPP vengono eluiti in condizioni di denaturazione. I campioni vengono preparati per l’analisi della spettrometria di massa mediante la rimozione di detergenti attraverso l’arricchimento proteico SP3, la digestione triptica e la desalinizzazione. I campioni possono quindi essere facoltativamente etichettati TMT prima dell’analisi della spettrometria di massa. Fare clic qui per visualizzare una versione più grande di questa figura.
PIB-MS comporta lisi e chiarificazione di cellule o tessuti, incubazione del lisato con PIB, eluizione e analisi dell’eluato tramite western blotting o approcci basati sulla spettrometria di massa (Figura 1). L’aggiunta di MCLR libero può essere utilizzata come controllo per distinguere specifici leganti PIB da interattori non specifici. Per la maggior parte delle applicazioni, un approccio senza etichette può essere utilizzato per identificare direttamente le proteine negli eluati. Nei casi in cui è necessaria una maggiore precisione nella quantificazione o nell’identificazione di specie a bassa abbondanza, è possibile utilizzare un’ulteriore elaborazione con etichettatura TMT (Tandem Mass-Tag) per aumentare la copertura e ridurre l’input.
