Proteinfosforylering kontrollerer de fleste cellulære prosesser, inkludert, men ikke begrenset til, responsen på DNA-skade, vekstfaktorsignalering og passasjen gjennom mitose 1,2,3. I pattedyrceller er flertallet av proteiner fosforylert ved en eller flere serin-, treon- eller tyrosinrester på et tidspunkt, med fosforeriner og fosforetoniner som består av omtrent 98% av alle fosforyleringssteder 2,3. Mens kinases har blitt grundig studert i cellulær signalering, vises fortsatt PPPs rolle i reguleringen av dynamiske cellulære prosesser.
Fosforyleringsdynamikken styres av det dynamiske samspillet mellom kinaser og fosfater. I pattedyrceller er det mer enn 400 proteinkinaser som katalyserer serin / treoninfosforylering. Over 90% av disse nettstedene er defosforylatert av fosfoproteinfosfater (PPPer), en liten familie av enzymer som består av PP1, PP2A, PP2B, PP4-7, PPT og PPZ 2,3. PP1 og PP2A er ansvarlige for de fleste fosfor og fosforondefosforylering i en celle 2,3,4. Den bemerkelsesverdige forskjellen i antall mellom kinases og fosfater og mangelen på spesifisitet av PPP katalytiske subenheter in vitro førte til troen på at kinases er den viktigste determinanten for fosforylering 2,3. Imidlertid har flere studier vist fosfater for å etablere substratspesifikkitet gjennom dannelsen av multimeriske holoenzymer 5,6,7,8,9. For eksempel er PP1 en heterodimer som består av en katalytisk underenhet og på et gitt tidspunkt en av de mer enn 150 regulatoriske underenhetene 6,7,8. På den annen side er PP2A en heterotrimer som er dannet av et stillas (A), en regulatorisk (B) og en katalytisk (C) underenhet 2,3,9. Det er fire forskjellige familier av PP2A regulatoriske underenheter (B55, B56, PR72 og striatin), hver med flere gener, skjøtevarianter og lokaliseringsmønstre 2,3,9. Den multimeriske naturen til PPPer fyller gapet i antall kinases og PPP katalytiske underenheter. Det skaper imidlertid analytiske utfordringer for å studere PPP-signalering. For å analysere PPP-signalering grundig, er det viktig å undersøke de forskjellige holoenzymene i en celle eller et vev. Store fremskritt har blitt gjort i å studere den menneskelige kinome gjennom bruk av kinase inhibitor perler, betegnet multiplex inhibitor perler eller kinobeads, en kjemisk proteomisk strategi der kinase inhibitors er immobilisert på perler og masse spektrometri brukes til å identifisere beriket kinases og deres interagere 10,11,12,13.
Vi har etablert en lignende tilnærming for å studere PPP-biologi. Denne teknikken innebærer affinitetsfangst av PPP-katalytiske underenheter ved hjelp av perler med en immobilisert, ikke-selektiv PPP-hemmer kalt microcystin-LR (MCLR) kalt fosfatasehemmerperler (PIB)14,15. I motsetning til andre metoder som krever endogen merking eller uttrykk for eksogene PPP-underenheter som kan endre proteinaktivitet eller lokalisering, tillater PIB-MS berikelse av endogene PPP-katalytiske underenheter, deres tilhørende regulatoriske og stillasunderenheter, og interagerende proteiner (kalt PPPome) fra celler og vev på et gitt tidspunkt eller under spesifikke behandlingsforhold. MCLR hemmer PP1, PP2A, PP4-6, PPT og PPZ ved nanomolarkonsentrasjoner, noe som gjør PIBer svært effektive til å berike for PPPome16. Denne metoden kan skaleres for bruk på ethvert startmateriale fra celler til kliniske prøver. Her beskriver vi i detalj bruken av PIBer og massespektrometri (PIB-MS) for effektivt å fange, identifisere og kvantifisere den endogene PPPome og dens modifikasjonstilstander.
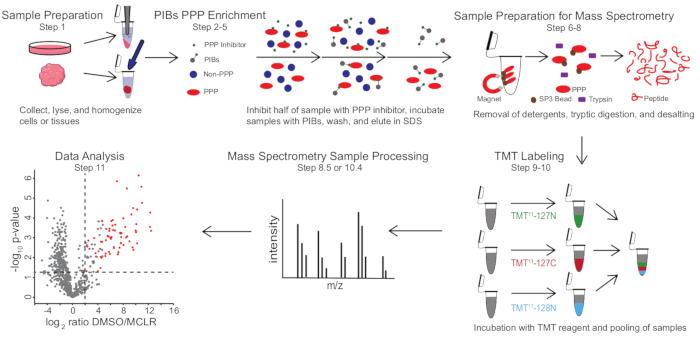
Figur 1: Visuelt sammendrag av PIB-MS-protokollen. I et PIB-MS-eksperiment kan prøver fås i forskjellige former, fra celler til svulster. Prøven samles inn, lyser og homogeniseres før PPP-berikelse. For å berike for PPPer inkuberes lysatet med PIBer med eller uten PPP-hemmer, for eksempel MCLR. PIB-ene vaskes deretter, og PPPer elutes under denatureringsforhold. Prøvene er utarbeidet for massespektrometrianalyse ved fjerning av vaskemidler gjennom SP3 proteinberikelse, tryptisk fordøyelse og avsalting. Prøver kan deretter eventuelt TMT-merket før massespektrometrianalyse. Klikk her for å se en større versjon av denne figuren.
PIB-MS innebærer lysis og avklaring av celler eller vev, inkubasjon av lysatet med PIBer, elution og analyse av eluate via vestlig blotting eller massespektrometribaserte tilnærminger (figur 1). Tillegg av gratis MCLR kan brukes som en kontroll for å skille spesifikke PIB-permer fra ikke-spesifikke interagere. For de fleste applikasjoner kan en etikettfri tilnærming brukes til å identifisere proteiner direkte i eluates. I tilfeller der større presisjon i kvantifisering eller identifisering av lav-overflod arter er nødvendig, kan videre behandling med tandem masse-tag (TMT) merking brukes til å øke dekningen og redusere inngang.
