Фосфорилирование белка контролирует большинство клеточных процессов, включая, но не ограничиваясь, реакцией на повреждение ДНК, передачей сигналов фактора роста и прохождением через митоз 1,2,3. В клетках млекопитающих большинство белков фосфорилируются в одном или нескольких остатках серина, треонина или тирозина в какой-то момент времени, причем фосфосерины и фосфотреонины составляют примерно 98% всех участков фосфорилирования 2,3. Хотя киназы широко изучаются в клеточной сигнализации, роль ПГЧС в регуляции динамических клеточных процессов все еще вырисовывается.
Динамика фосфорилирования контролируется динамическим взаимодействием между киназами и фосфатазами. В клетках млекопитающих насчитывается более 400 протеинкиназ, которые катализируют фосфорилирование серина/треонина. Более 90% этих участков дефосфорилируются фосфопротеинфосфатазами (ППС), небольшим семейством ферментов, которое состоит из PP1, PP2A, PP2B, PP4-7, PPT и PPZ 2,3. PP1 и PP2A ответственны за большую часть дефосфосерина и фосфотреонина в клетке 2,3,4. Заметная разница в количестве между киназами и фосфатазами и отсутствие специфичности каталитических субъединиц ППС in vitro привели к убеждению, что киназы являются основным детерминантом фосфорилирования 2,3. Однако многочисленные исследования показали, что фосфатазы устанавливают субстратную специфичность за счет образования многомерных голоферментов 5,6,7,8,9. Например, PP1 представляет собой гетеродимер, который состоит из каталитической субъединицы и, в данный момент времени, одной из более чем 150 регуляторных субъединиц 6,7,8. И наоборот, PP2A представляет собой гетеротример, который образован из строительных лесов (A), регулятора (B) и каталитической (C) субъединицы 2,3,9. Существует четыре различных семейства регуляторных субъединиц PP2A (B55, B56, PR72 и стриатин), каждое из которых имеет несколько генов, вариантов сращивания и паттернов локализации 2,3,9. Многомерный характер ПГЧС заполняет пробел в количестве киназ и каталитических субъединиц ППС. Однако это создает аналитические проблемы для изучения сигнализации ГЧП. Чтобы всесторонне проанализировать передачу сигналов ППС, крайне важно исследовать различные голоферменты в клетке или ткани. Большие успехи были достигнуты в изучении человеческого кинома благодаря использованию шариков ингибитора киназы, называемых бусинами мультиплексных ингибиторов или кинобеодами, химической протеомной стратегии, в которой ингибиторы киназы иммобилизуются на шариках, а масс-спектрометрия используется для идентификации обогащенных киназ и их интеракторов 10,11,12,13.
Мы установили аналогичный подход к изучению биологии ГЧП. Этот метод включает в себя захват аффинности каталитических субъединиц ППС с использованием шариков с иммобилизованным, неселективным ингибитором ППС, называемым микроцистином-LR (MCLR), называемым шариками ингибитора фосфатазы (PIBs)14,15. В отличие от других способов, требующих эндогенной маркировки или экспрессии экзогенных субъединиц ППС, которые могут изменять активность или локализацию белка, PIB-MS позволяет обогащать эндогенные каталитические субъединицы ППС, связанные с ними регуляторные и каркасные субъединицы, а также взаимодействующие белки (называемые PPPome) из клеток и тканей в данный момент времени или при определенных условиях обработки. MCLR ингибирует PP1, PP2A, PP4-6, PPT и PPZ в наномолярных концентрациях, что делает PIB высокоэффективными при обогащении PPPome16. Этот метод может быть масштабирован для использования на любом исходном материале от клеток до клинических образцов. Здесь мы подробно описываем использование PIB и масс-спектрометрии (PIB-MS) для эффективного захвата, идентификации и количественной оценки эндогенного PPPome и его модифицирующих состояний.
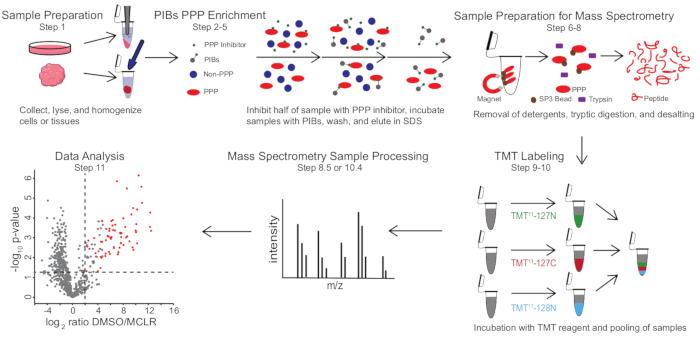
Рисунок 1: Визуальная сводка протокола PIB-MS. В эксперименте PIB-MS образцы могут быть получены в различных формах, от клеток до опухолей. Образец собирается, лизируется и гомогенизируется перед обогащением ППС. Для обогащения для ПГЧС лизат инкубируют с ПИБ с ингибитором ППС или без него, таким как MCLR. Затем ПИБ промывают, а ПГЧС элюируют в условиях денатурации. Образцы подготавливаются для масс-спектрометрического анализа путем удаления моющих средств путем обогащения белка SP3, триптического сбраживания и обессоливания. Затем образцы могут быть опционально помечены TMT перед масс-спектрометрическим анализом. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
PIB-MS включает лизис и осветление клеток или тканей, инкубацию лизата с PIB, элюирование и анализ элюата с помощью подходов, основанных на вестерн-блоттинге или масс-спектрометрии (рисунок 1). Добавление свободного MCLR может быть использовано в качестве элемента управления для различения конкретных связующих PIB от неспецифических интеракторов. Для большинства применений подход без маркировки может быть использован для непосредственной идентификации белков в элюатах. В тех случаях, когда требуется более высокая точность количественной оценки или идентификация видов с низкой численностью, для увеличения охвата и уменьшения вводимых ресурсов может использоваться дальнейшая обработка с помощью маркировки тандемной массы (ТМТ).
