La fosforilación de proteínas controla la mayoría de los procesos celulares, incluyendo pero no limitado a la respuesta al daño del ADN, la señalización del factor de crecimiento y el paso a través de la mitosis 1,2,3. En las células de mamíferos, la mayoría de las proteínas se fosforilan en uno o más residuos de serina, treonina o tirosina en algún momento, con fosfoserinas y fosfotreoninas que comprenden aproximadamente el 98% de todos los sitios de fosforilación 2,3. Si bien las quinasas se han estudiado ampliamente en la señalización celular, el papel de las APP en la regulación de los procesos celulares dinámicos aún está emergiendo.
La dinámica de fosforilación está controlada por la interacción dinámica entre las quinasas y las fosfatasas. En las células de mamíferos, hay más de 400 proteínas quinasas que catalizan la fosforilación de serina/treonina. Más del 90% de estos sitios están desfosforilados por fosfoproteínas fosfatasas (APP), una pequeña familia de enzimas que consiste en PP1, PP2A, PP2B, PP4-7, PPT y PPZ 2,3. PP1 y PP2A son responsables de la mayoría de la desfosforilación de fosfoserina y fosfotreonina dentro de una célula 2,3,4. La notable diferencia de número entre quinasas y fosfatasas y la falta de especificidad de las subunidades catalíticas de PPP in vitro llevaron a la creencia de que las quinasas son el principal determinante de la fosforilación 2,3. Sin embargo, múltiples estudios han demostrado que las fosfatasas establecen la especificidad del sustrato a través de la formación de holoenzimas multiméricas 5,6,7,8,9. Por ejemplo, PP1 es un heterodímero que consiste en una subunidad catalítica y, en un momento dado, una de las más de 150 subunidades reguladoras 6,7,8. Por el contrario, PP2A es un heterotrímero que está formado por un andamio (A), un regulador (B) y una subunidad catalítica (C) 2,3,9. Hay cuatro familias distintas de subunidades reguladoras de PP2A (B55, B56, PR72 y estriatina), cada una con múltiples genes, variantes de empalme y patrones de localización 2,3,9. La naturaleza multimérica de las APP llena el vacío en el número de quinasas y subunidades catalíticas de PPP. Sin embargo, crea desafíos analíticos para estudiar la señalización de PPP. Para analizar exhaustivamente la señalización de PPP, es fundamental investigar las diversas holoenzimas dentro de una célula o tejido. Se han logrado grandes avances en el estudio del quinasmo humano mediante el uso de perlas inhibidoras de la quinasa, denominadas perlas inhibidoras multiplex o quinabiadas, una estrategia proteómica química donde los inhibidores de la quinasa se inmovilizan en perlas y se utiliza la espectrometría de masas para identificar quinasas enriquecidas y sus interactores 10,11,12,13.
Hemos establecido un enfoque similar para estudiar la biología de la PPP. Esta técnica consiste en la captura por afinidad de subunidades catalíticas de PPP utilizando perlas con un inhibidor de PPP inmovilizado y no selectivo llamado microcistina-LR (MCLR) denominado perlas inhibidoras de la fosfatasa (PIBs)14,15. A diferencia de otros métodos que requieren el etiquetado endógeno o la expresión de subunidades exógenas de PPP que podrían alterar la actividad o localización de proteínas, PIB-MS permite el enriquecimiento de subunidades catalíticas endógenas de PPP, sus subunidades reguladoras y de andamiaje asociadas, y proteínas que interactúan (denominadas PPPome) de células y tejidos en un punto de tiempo dado o bajo condiciones de tratamiento específicas. MCLR inhibe PP1, PP2A, PP4-6, PPT y PPZ a concentraciones nanomolares, lo que hace que los PIB sean altamente efectivos para enriquecer el PPPome16. Este método se puede escalar para su uso en cualquier material de partida, desde células hasta muestras clínicas. Aquí, describimos en detalle el uso de PIBs y espectrometría de masas (PIB-MS) para capturar, identificar y cuantificar eficientemente el PPPome endógeno y sus estados de modificación.
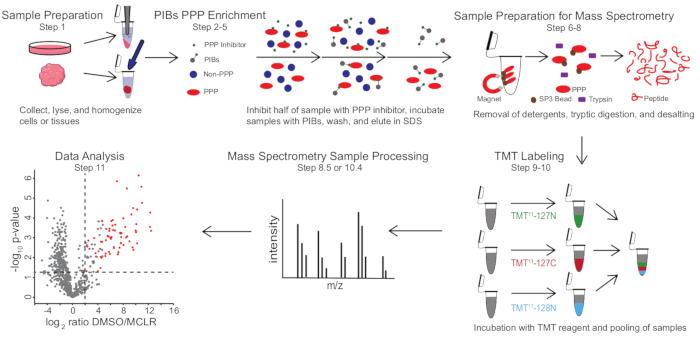
Figura 1: Resumen visual del protocolo PIB-MS. En un experimento PIB-MS, se pueden obtener muestras en varias formas, desde células hasta tumores. La muestra se recoge, se lisa y homogeneiza antes del enriquecimiento con PPP. Para enriquecer para las APP, el lisado se incuba con PIBs con o sin un inhibidor de PPP, como MCLR. Los PIB se lavan y las APP se eluyen en condiciones de desnaturalización. Las muestras se preparan para el análisis de espectrometría de masas mediante la eliminación de detergentes a través del enriquecimiento de proteínas SP3, la digestión tríptica y la desalinización. Las muestras pueden ser opcionalmente marcadas con TMT antes del análisis de espectrometría de masas. Haga clic aquí para ver una versión más grande de esta figura.
PIB-MS implica lisis y clarificación de células o tejidos, incubación del lisado con PIBs, elución y análisis del eluido a través de western blotting o enfoques basados en espectrometría de masas (Figura 1). La adición de MCLR libre se puede utilizar como un control para distinguir aglutinantes ESPECÍFICOS de PIB de interactores no específicos. Para la mayoría de las aplicaciones, se puede utilizar un enfoque sin etiquetas para identificar directamente las proteínas en los eluidos. En los casos en que se necesita una mayor precisión en la cuantificación o la identificación de especies de baja abundancia, se puede utilizar un procesamiento adicional con etiquetado de etiqueta de masa en tándem (TMT) para aumentar la cobertura y disminuir la entrada.
