Protein fosforilasyonu, DNA hasarına verilen yanıt, büyüme faktörü sinyallemesi ve mitoz 1,2,3’ten geçiş dahil ancak bunlarla sınırlı olmamak üzere çoğu hücresel süreci kontrol eder. Memeli hücrelerinde, proteinlerin çoğunluğu bir veya daha fazla serin, treonin veya tirozin kalıntısında fosforile edilir, fosfoserlerin ve fosfotreoninler tüm fosforilasyon bölgelerinin yaklaşık% 98’ini oluşturur 2,3. Kinazlar hücresel sinyalizasyonda kapsamlı bir şekilde incelenmiş olsa da, PPP’lerin dinamik hücresel süreçlerin düzenlenmesindeki rolü hala ortaya çıkmaktadır.
Fosforilasyon dinamikleri, kinazlar ve fosfatazlar arasındaki dinamik etkileşim ile kontrol edilir. Memeli hücrelerinde, serin / treonin fosforilasyonunu katalize eden 400’den fazla protein kinaz vardır. Bu bölgelerin% 90’ından fazlası, PP1, PP2A, PP2B, PP4-7,PPT ve PPZ 2,3’ten oluşan küçük bir enzim ailesi olan fosfoprotein fosfatazlar (PPP’ler) tarafından defosforile edilir. PP1 ve PP2A, bir hücre 2,3,4 içindeki fosfoserin ve fosfotreonin defosforilasyonunun çoğundan sorumludur. Kinazlar ve fosfatazlar arasındaki kayda değer sayı farkı ve PPP katalitik alt birimlerinin in vitro özgüllüğünün olmaması, kinazların fosforilasyonun ana belirleyicisi olduğu inancına yol açmıştır 2,3. Bununla birlikte, birçok çalışma, fosfatazların multimerik holoenzimlerinoluşumu yoluyla substrat özgüllüğünü oluşturduğunu göstermiştir 5,6,7,8,9. Örneğin, PP1, katalitik bir alt birimden ve belirli bir zamanda, 150’den fazla düzenleyici alt birim 6,7,8’den birinden oluşan bir heterodimerdir. Tersine, PP2A, bir iskele (A), bir düzenleyici (B) ve bir katalitik (C) alt birimi 2,3,9’dan oluşan bir heterotrimerdir. PP2A düzenleyici alt birimlerinin (B55, B56, PR72 ve striatin) dört ayrı ailesi vardır, her biri birden fazla gen, ekleme varyantı ve lokalizasyon paternleri 2,3,9’dur. PPP’lerin multimerik doğası, kinazların ve PPP katalitik alt birimlerinin sayısındaki boşluğu doldurur. Bununla birlikte, PPP sinyallemesini incelemek için analitik zorluklar yaratır. PPP sinyalini kapsamlı bir şekilde analiz etmek için, bir hücre veya doku içindeki çeşitli holoenzimleri araştırmak çok önemlidir. İnsan kinomunun incelenmesinde, multipleks inhibitör boncuklar veya kinoboncuklar olarak adlandırılan kinaz inhibitörü boncukların kullanılmasında, kinaz inhibitörlerinin boncuklar üzerinde hareketsiz hale getirildiği ve zenginleştirilmiş kinazları ve bunların interaktörlerini tanımlamak için kütle spektrometrisinin kullanıldığı kimyasal bir proteomik strateji kullanılarak büyük ilerlemeler kaydedilmiştir10,11,12,13.
PPP biyolojisini incelemek için benzer bir yaklaşım oluşturduk. Bu teknik, fosfataz inhibitörü boncuklar (PIB’ler) olarak adlandırılan mikrosistin-LR (MCLR) adı verilen hareketsiz, seçici olmayan bir PPP inhibitörü ile boncuklar kullanılarak PPP katalitik alt birimlerinin afinite yakalamasını içerir14,15. Protein aktivitesini veya lokalizasyonunu değiştirebilecek eksojen PPP alt birimlerinin endojen etiketlemesini veya ekspresyonunu gerektiren diğer yöntemlerden farklı olarak, PIB-MS, endojen PPP katalitik alt birimlerinin, bunlarla ilişkili düzenleyici ve iskele alt birimlerinin ve belirli bir zaman noktasında veya belirli tedavi koşulları altında hücrelerden ve dokulardan etkileşime giren proteinlerin (PPPome olarak adlandırılır) zenginleştirilmesine izin verir. MCLR, nanomolar konsantrasyonlarda PP1, PP2A, PP4-6, PPT ve PPZ’yi inhibe ederek PIB’leri PPPome16 için zenginleştirmede oldukça etkili hale getirir. Bu yöntem, hücrelerden klinik örneklere kadar herhangi bir başlangıç materyalinde kullanılmak üzere ölçeklendirilebilir. Burada, endojen PPPome ve modifikasyon durumlarını verimli bir şekilde yakalamak, tanımlamak ve ölçmek için PIB’lerin ve kütle spektrometresinin (PIB-MS) kullanımını ayrıntılı olarak açıklıyoruz.
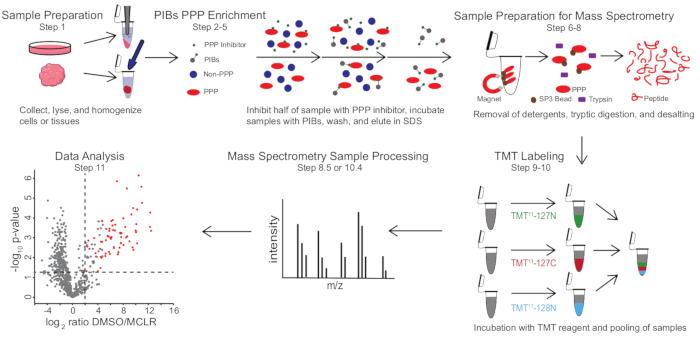
Şekil 1: PIB-MS protokolünün görsel özeti. Bir PIB-MS deneyinde, hücrelerden tümörlere kadar çeşitli şekillerde örnekler elde edilebilir. Numune, PPP zenginleştirmeden önce toplanır, lize edilir ve homojenize edilir. PPP’ler için zenginleştirmek için, lizat, MCLR gibi bir PPP inhibitörü olan veya olmayan PIB’lerle inkübe edilir. PIB’ler daha sonra yıkanır ve PPP’ler denatüre koşullarında salınır. Numuneler, SP3 protein zenginleştirme, triptik sindirim ve tuzdan arındırma yoluyla deterjanların uzaklaştırılması yoluyla kütle spektrometresi analizi için hazırlanır. Numuneler daha sonra kütle spektrometresi analizinden önce isteğe bağlı olarak TMT etiketli olabilir. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
PIB-MS, hücrelerin veya dokuların lizisini ve berraklaştırılmasını, lizatın PIB’lerle inkübasyonunu, elüsyonu ve batı lekelenmesi veya kütle spektrometrisine dayalı yaklaşımlarla elüatın analizini içerir (Şekil 1). Serbest MCLR’nin eklenmesi, spesifik PIB bağlayıcılarını spesifik olmayan interaktörlerden ayırt etmek için bir kontrol olarak kullanılabilir. Çoğu uygulama için, elüatlardaki proteinleri doğrudan tanımlamak için etiketsiz bir yaklaşım kullanılabilir. Nicelemede veya düşük bolluktaki türlerin tanımlanmasında daha fazla hassasiyetin gerekli olduğu durumlarda, kapsama alanını artırmak ve girişi azaltmak için tandem kütle etiketi (TMT) etiketleme ile daha fazla işlem kullanılabilir.
