The human body harbors an estimated 10-100 trillion live microbial cells (bacteria, archaea fungi), which are primarily found in the gut, skin, and mucosal environments1. In a healthy state, these provide benefits to their host, including vitamin production, maturation of the immune system, stimulation of innate and adaptive immune responses to pathogens, regulation of fat metabolism, modulation of stress responses, and more, with an impact on growth and development, disease onset, and ageing2,3,4,5. The gut microbiota also evolves considerably throughout life. The most drastic evolution occurs during infancy and early childhood6, but significant changes also occur with age, including a decrease in Bifidobacterium abundance and an increase in Clostridium, Lactobacillus, Enterobacteriaceae, and Enterococcus species7. Lifestyle can further alter gut microbial composition leading to dysbiosis (loss of beneficial bacteria, overgrowth of opportunistic bacteria), resulting in various pathologies such as inflammatory bowel disease, diabetes, and obesity5, but also contributing to Alzheimer's and Parkinson's diseases8,9,10,11.
This realization has critically contributed to refining the concept of the gut-brain axis (GBA), where interactions between gut physiology (now including the microbes within it) and the nervous system are considered the main regulator of animal metabolism and physiological functions12. However, the precise role of microbiota in gut-brain signaling and the associated mechanisms of action are far from being fully understood13. With gut microbiota being a key determinant of healthy aging, how bacteria modulate the aging process has become a subject of intense research and controversy6,14,15.
With the demonstration that the roundworm Caenorhabditis elegans hosts a bonafide gut microbiota dominated-as in other species-by Bacteroidetes, Firmicutes, and Actinobacteria16,17,18,19,20, its rapid rise as an experimental platform to study host-gut commensal interactions21,22,23,24,25,26 has significantly expanded our investigative arsenal26,27,28,29. In particular, high-throughput experimental approaches available for C. elegans to study gene-diet, gene-drug, gene-pathogen, etc. interactions, can be adapted to rapidly explore how bacterial isolates and cocktails impact C. elegans health and aging.
The present protocol describes an experimental pipeline to screen at once arrays of bacterial isolates or mixtures set in multiwell plates for effects on C. elegans stress resistance as a proxy for health, which can be used to identify probiotics. It details how to grow large worm populations and handle bacterial arrays in 96- and 384-well plate formats before processing worms for automated stress resistance analysis using a fluorescence plate reader (Figure 1). The approach is based on label-free automated survival assays (LFASS)30 that exploit the phenomenon of death fluorescence31, whereby dying worms produce a burst of blue fluorescence that can be used to pinpoint the time of death. Blue fluorescence is emitted by glucosyl esters of anthranilic acid stored in C. elegans gut granules (a type of lysosome-related organelle), which burst when a necrotic cascade is triggered in the worm gut upon death31.
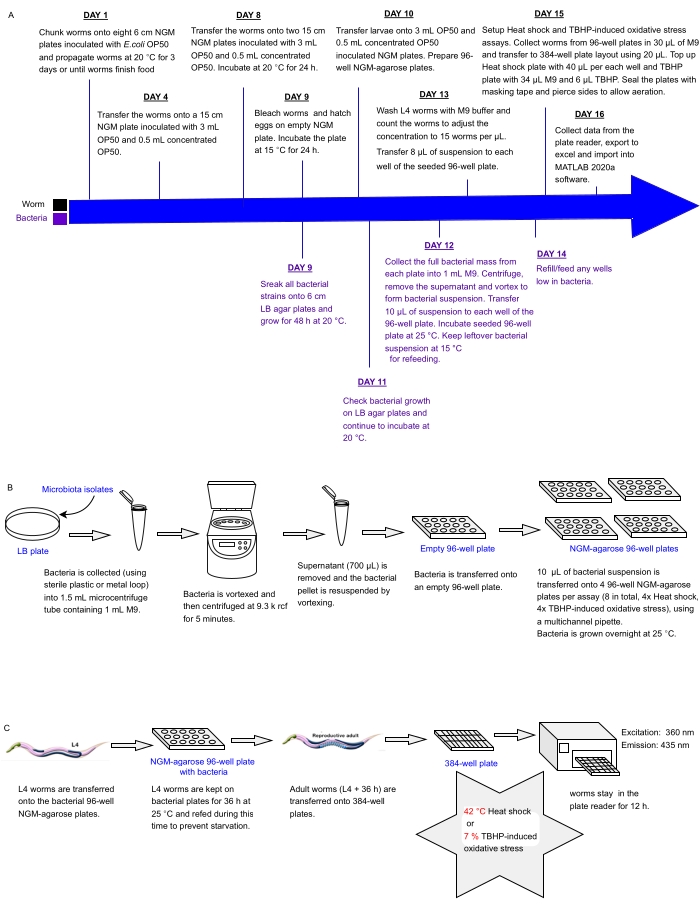
Figure 1: Experimental workflow for high-throughput screening of bacterial isolates with impact on C. elegans resistance to stress. (A) Timeline for worm and bacterial maintenance and assay setup. (B) 96-well bacterial plate array setup and handling. (C) 384-well worm plate setup. Please click here to view a larger version of this figure.
