İnsan vücudu, öncelikle bağırsak, cilt ve mukozal ortamlarda bulunan tahmini 10-100 trilyon canlı mikrobiyal hücreyi (bakteri, arkea mantarı) barındırır1. Sağlıklı bir durumda, bunlar vitamin üretimi, bağışıklık sisteminin olgunlaşması, patojenlere karşı doğuştan gelen ve adaptif bağışıklık tepkilerinin uyarılması, yağ metabolizmasının düzenlenmesi, stres yanıtlarının modülasyonu ve daha fazlası dahil olmak üzere konakçılarına büyüme ve gelişme, hastalık başlangıcı ve yaşlanma üzerinde etkili olan faydalar sağlar 2,3,4,5 . Bağırsak mikrobiyotası da yaşam boyunca önemli ölçüde gelişir. En şiddetli evrim bebeklik ve erken çocuklukdöneminde meydana gelir 6, ancak Bifidobacterium bolluğunda bir azalma ve Clostridium, Lactobacillus, Enterobacteriaceae ve Enterococcus türlerinde bir artış da dahil olmak üzere yaşla birlikte önemli değişiklikler meydana gelir7. Yaşam tarzı, dysbiosis’e (faydalı bakteri kaybı, fırsatçı bakterilerin aşırı büyümesi) yol açan bağırsak mikrobiyal bileşimini daha da değiştirebilir, enflamatuar bağırsak hastalığı, diyabet ve obezite5 gibi çeşitli patolojilere neden olabilir, ancak aynı zamanda Alzheimer ve Parkinson hastalıklarına da katkıda bulunabilir 8,9,10,11.
Bu gerçekleşme, bağırsak fizyolojisi (şimdi içindeki mikroplar dahil) ve sinir sistemi arasındaki etkileşimlerin hayvan metabolizmasının ve fizyolojik işlevlerin ana düzenleyicisi olarak kabul edildiği bağırsak-beyin ekseni (GBA) kavramının rafine edilmesine kritik katkıda bulunmuştur12. Bununla birlikte, mikrobiyotanın bağırsak-beyin sinyalizasyonundaki kesin rolü ve ilişkili etki mekanizmaları tam olarak anlaşılmaktan uzaktır13. Bağırsak mikrobiyotasının sağlıklı yaşlanmanın önemli bir belirleyicisi olmasıyla, bakterilerin yaşlanma sürecini nasıl modüle ettiği yoğun bir araştırma ve tartışma konusu haline gelmiştir 6,14,15.
Yuvarlak kurt Caenorhabditis elegans’ın, diğer türlerde olduğu gibi, Bacteroidetes, Firmicutes ve Actinobacteria16,17,18,19,20 tarafından domine edilen iyi niyetli bir bağırsak mikrobiyotasına ev sahipliği yaptığının gösterilmesiyle, konakçı-bağırsak kommensal etkileşimlerini incelemek için deneysel bir platform olarak hızlı yükselişi21,22,23,24 ,25,26 araştırma cephaneliğimizi önemli ölçüde genişletti26,27,28,29. Özellikle, C. elegans’ın gen-diyet, gen-ilaç, gen-patojen vb. etkileşimlerini incelemesi için mevcut olan yüksek verimli deneysel yaklaşımlar, bakteriyel izolatların ve kokteyllerin C. elegans sağlığını ve yaşlanmasını nasıl etkilediğini hızlı bir şekilde keşfetmek için uyarlanabilir.
Mevcut protokol, probiyotikleri tanımlamak için kullanılabilecek sağlık için bir vekil olarak C. elegans stres direnci üzerindeki etkileri için çok kuyulu plakalara yerleştirilmiş bakteri izolatlarının veya karışımlarının dizilerini bir kerede taramak için deneysel bir boru hattını tanımlamaktadır. Bir floresan plaka okuyucu kullanarak otomatik stres direnci analizi için solucanları işlemeden önce büyük solucan popülasyonlarının nasıl yetiştirileceğini ve bakteri dizilerinin 96 ve 384 kuyucuklu plaka formatlarında nasıl ele alınacağını detaylandırır (Şekil 1). Yaklaşım, ölüm floresansı31 fenomeninden yararlanan etiketsiz otomatik hayatta kalma tahlillerine (LFASS)30 dayanmaktadır; burada ölmekte olan solucanlar, ölüm zamanını belirlemek için kullanılabilecek bir mavi floresan patlaması üretir. Mavi floresan, C. elegans bağırsak granüllerinde (lizozom ile ilişkili bir tür organel) depolanan antranilik asidin glukozil esterleri tarafından yayılır ve bu da ölüm üzerine solucan bağırsağında nekrotik bir kaskad tetiklendiğinde patlar31.
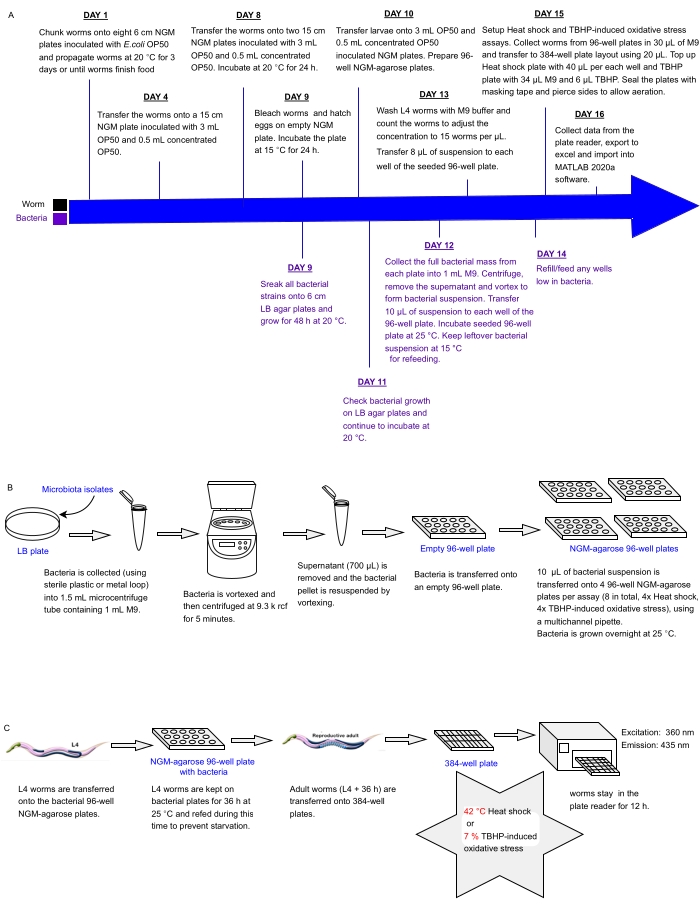
Şekil 1: C. elegans’ın strese karşı direnci üzerinde etkisi olan bakteriyel izolatların yüksek verimli taranması için deneysel iş akışı . (A) Solucan ve bakteri bakımı ve tahlil kurulumu için zaman çizelgesi. (B) 96 kuyucuklu bakteri plakası dizisi kurulumu ve kullanımı. (C) 384 delikli sonsuz plaka kurulumu. Bu şeklin daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
