Electroporation to facilitate gene transfer in skeletal muscle is a useful technique used to evaluate changes in muscle physiology. We have demonstrated a detailed, step-by-step procedure to accomplish efficient gene transfer in both the TA and EDL muscles. Differences in transfection efficiency occur due to a number of variables. Among these variables are electroporation parameters (pulses, voltage, pulse duration, etc.), gene construct size, and concentration/volume of DNA injected. We have previously shown that electroporation parameters of 5 pulses at 125 V/cm, with 20 ms duration separated by 200 ms intervals, are sufficient to accomplish efficient gene transfer in the TA14. We also demonstrate in the current study that injection/electroporation of DNA does not cause loss of muscle contractility in the EDL 3-days post-experimentation.
In order to visualize gene transfer, a pcDNA3-EGFP or pcDNA3 (control) construct were electroporated into mouse TA or EDL muscle. 3 days after the procedure, the TA or EDL was carefully dissected, placed in optimal cutting temperature (OTC) media, and snap-frozen in liquid nitrogen-cooled isopentane (2-methylbutane), as previously described14,17,18,19. 10 µm muscle sections were obtained using a cryostat taken from the mid-belly of each muscle. Sections were then incubated in 1% paraformaldehyde for 5 min, followed by 3-5 min washes in PBS. Sections were then incubated in wheat-germ agglutinin conjugated to Texas Red diluted 1:100 in PBS for 90 min in the dark. Sections were again washed in PBS 3 times for 5 min each. Muscle sections were then coverslipped in aqueous mounting media and imaged in the 594 nm wavelength (red) to visualize the individual muscle fibers and in 480 nm wavelength to detect GFP (Figure 1A). The control muscles injected with pcDNA3 were imaged in both channels with the same exposure settings to control for potential autofluorescence. Muscles injected/electroporated with EGFP showed positive green fibers, demonstrating uptake of the pcDNA3-EGFP construct, while muscles injected/electroporated with pcDNA3 (control) showed no green positive fibers. We and others have previously shown that myofiber cross-sectional area is not compromised by GFP expression by comparing GFP positive fibers (green) to non-GFP expressing fibers (black) within the same muscle sections4,6,19. When imaged at a lower magnification, EDL muscle transfection efficiency is visualized through the appearance of green fibers (positive) versus black fibers (negative) (Figure 1B). Transfection efficiency using this procedure was 56.6% ± 4.7%, as measured in 3 EDL muscles. This data shows that the injection and electroporation of the TA muscle is sufficient for efficient gene transfer. Additionally, injection and electroporation of the EDL elicits efficient uptake of gene constructs.
An important tool for the evaluation of skeletal muscle physiology is the measurement of muscle contractility. Previous investigators have shown that the contractility of the TA measured in situ may be compromised early after injection and electroporation of the hind limb13. In order to test whether EDL contractility is compromised after injection and electroporation, we surgically exposed the EDL, injected it with pcDNA3-EGFP or construct, and electroporated the hindlimb. As a control, the alternate limb was left untouched for comparison. The mice were euthanized 3 days later, and the EDL whole muscle contractility was measured using field stimulation in a physiological bath, as previously described14,19,20,21. Briefly, the EDL was dissected, and the tendons were tied via 4/0 silk suture to stainless steel hooks and suspended between a force transducer and static base. The muscles were bathed in Tyrode buffer, which is a physiological bath solution (121 mM NaCl, 5.0 mM KCl, 1.8 mM CaCl2, 0.5 mM MgCl2, 0.4 mM NaH2PO4, 24 mM NaHCO3, 0.1 mM EDTA, 5.5 mM glucose) and bubbled with 100% oxygen throughout the procedure. Muscle contraction was stimulated using platinum electrodes to deliver a supramaximal stimulus. Muscle optimal length (Lo) was adjusted to yield the maximum force at the beginning of the procedure. The force-frequency relationship was determined using stimulation frequencies between 1-150 Hz (0.5 ms pulses at supramaximal voltage). The muscle was allowed to relax for 3 min between each stimulation. After the stimulation procedure was completed, the weight and length of the muscle were measured. The specific force was calculated by normalizing the absolute force to the whole muscle cross-sectional area, which is calculated as the weight divided by the length and using the previously determined muscle density constant (1.056 kg/m−3)21. We have shown that representative injected and electroporated EDLs have similar tetanic responses at 100 Hz compared to control non-injected or electroporated muscles (Figure 2A and Figure 2B). We found muscle tetanic force, specific tetanic force, time to peak tension, and half relaxation time were not compromised in the injected and electroporated EDL compared to the untouched control (Table 1). Our data demonstrate that protein expression that affects contractility may be modulated using electroporation in the EDL without causing adverse effects.
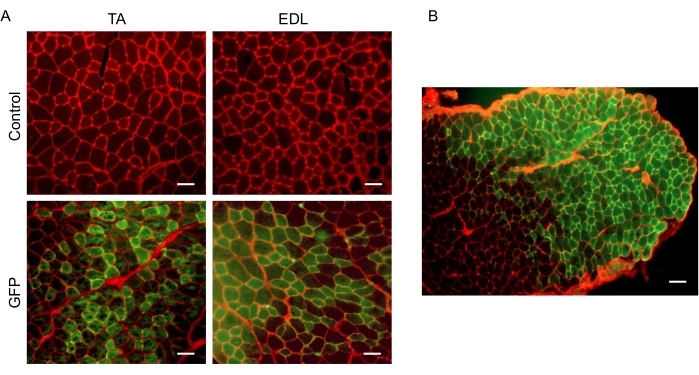
Figure 1: Electroporation of pcDNA3-EGFP is sufficient for DNA uptake in TA and EDL muscle. A) Representative cross-sections from TA and EDL controls (injected with control vector and electroporated 125 V/cm) and GFP (injected with pcDNA3-EGFP and electroporated 125 V/cm). Scale bar 50 µm. B) Representative cross-section from the EDL demonstrating transfection efficiency. Scale bar 100 µm. Please click here to view a larger version of this figure.
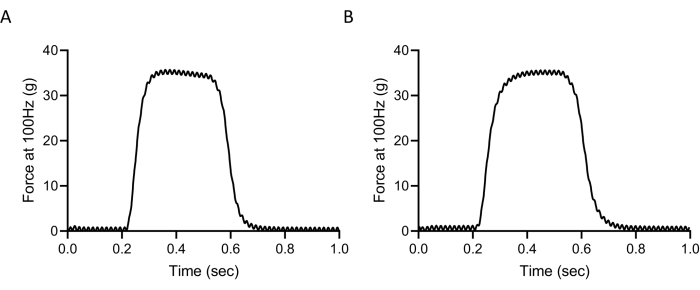
Figure 2: Injection and electroporation does not compromise muscle function. Representative tetanic force curves at 100 Hz from A) non-injected/electroporated EDL (control) and B) injected/electroporated EDL (GFP). Please click here to view a larger version of this figure.
| Control (n=3) | GFP (n=3) | p-value | |
| FO (g) | 34.13 ± 1.15 | 35.87 ± 1.55 | 0.1918 |
| sFO (kN/m2) | 691.56 ± 45.80 | 660.00 ± 33.61 | 0.3917 |
| TTP (sec) | 0.249 ± 0.0203 | 0.247 ± 0.0197 | 0.9084 |
| RT½ (sec) | 0.035 ± 0.0035 | 0.033 ± .0031 | 0.6458 |
| Control -non-injected/non-electroporated; GFP-injected with pcDNA3-EGFP and electroporated. Values are mean ± SD. p=0.05 considered significant. F0– Raw Tetanic Force at 100Hz, sF0-Tetanic Specific Force at 100Hz, TTP- Time to Peak at 1Hz, RT½- Time to half relaxation at 1Hz. | |||
Table 1: Whole muscle contractility from control versus electroporated EDLs. Table showing that different muscle force parameters are not compromised in injected/electroporated EDLs (GFP) compared to non-injected/non-electroporated controls. F0 = tetanic force at 100 Hz, sF0 = specific tetanic force at 100 Hz, TTP = time to peak tension at 1 Hz, RT½ = half relaxation time. Student's t-test; Significance at p = 0.05. n = 3.
