All experimental procedures were approved by the Malmö/Lund Animal Research Ethics Board, ethical permit number 12548-19, and conducted in agreement with the Swedish Animal Welfare Agency regulations and the EU Directive 2010/63/EU for animal experiments. C57BL6/J and Contactin-associated protein-like 2 (Cntnap2) knock-out (KO) mice, both males and females, were used at postnatal day (P) 2 for the present study. Human embryonic stem cells (hESCs) were used. The animals and stem cells were obtained from commercial sources (see Table of Materials).
1. Generation of the hdIN precursors
NOTE: All the steps in this section are done in a cell culture hood. The hESCs were maintained as feeder-free cells on coated plates using a stem cell culture medium and passaged as colonies.
- Perform forward programming of the hESCs to hdIN precursors (Figure 1).
NOTE: The forward programming approach is based on the procedure described in Gonzalez-Ramos et al.8 and Yang et al.9.- Transduce the hESCs with lentiviral vectors containing the Tet-On system in a fresh stem cell culture medium containing 10 µM of ROCK inhibitor (RI) (see Table of Materials). Keep at 37 °C in the incubator.
- Change the medium to a fresh stem cell culture medium 16 h after transduction and maintain the transduced cells in the same way as the non-transduced hESCs.
- When confluent, detach the hESCs as single cells using a cell detachment solution (see Table of Materials) and plate them in coated 6-well plates at a density of 3 x 105 cells/well in stem cell culture medium containing 10 µM of RI on day in vitro (DIV) −1.
- At 1 DIV, replace the culturing medium with N2 medium containing 2 g/L of doxycycline (DOX, see Table of Materials).
NOTE: DOX is used to induce transgene expression of Ascl1 and Dlx29, from 1 DIV to 7 DIV, and then continues in vivo by its addition to the drinking water25. - At 2 DIV, an antibiotic resistance selection period starts that will last until 5 DIV. Add puromycin (puro) and hygromycin (hygro) to the fresh medium (see Table of Materials). Change the medium on 2 DIV, 4 DIV, and 5 DIV.
NOTE: Optimize the concentrations of puro and hygro before the differentiation protocol using transduced and non-transduced hESCs to ensure the survival of only the cells carrying the antibiotic resistance cassettes. In this protocol, 0.5 µg/mL of puro and 750 µg/mL of hygro were used. - At 5 DIV, the antibiotic selection period ends. Change to N2 medium supplemented with 2 g/L of DOX and 4 µM of cytosine β-D-arabinofuranoside (Ara-C, see Table of Materials).
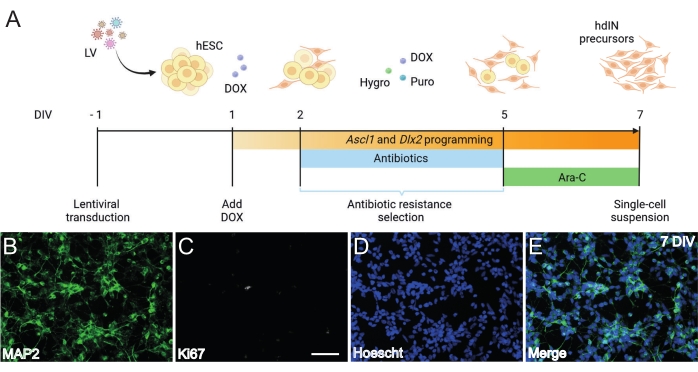
Figure 1: Generation of hdIN precursors from hESCs by overexpressing Ascl1 and Dlx2. (A) Schematics of the differentiation protocol used for the generation of hdIN precursors. (B–E) Immunocytochemistry of hdIN precursors at 7 DIV for (B) the neuronal marker MAP2, (C) the proliferative marker Ki67, (D) general nuclear staining, and (E) a merge of the previous markers. Scale bar: 50 µm. Please click here to view a larger version of this figure.
2. Preparation of the single-cell suspension for transplantation
NOTE: All the steps in this section are done in the cell culture hood. On 7 DIV, hdIN precursors are dissociated and used for transplantation.
- Perform detachment of the cells following the steps below.
- Remove the medium and carefully rinse the cells with DPBS without calcium and magnesium.
- Add 400 µL of the cell detachment solution (see Table of Materials) to each well of the 6-well plate.
NOTE: Ensure the coverage of the whole surface by the solution. - Incubate for 2-3 min at 37 °C in the incubator until the border of the cells starts to look shiny, indicating enzymatic degradation of cell surface proteins and detachment from the well surface.
NOTE: To increase cell survival, do not wait too long when all the cells are completely detached and floating around. - Then, add 600 µL of fresh N2 medium to each well of the 6-well plate to stop the enzyme, and mechanically with the pipette, help to detach the cells to obtain a single-cell suspension.
- Transfer the cell suspension to a plastic tube (15 mL) and centrifuge at 180 x g for 4 min at room temperature (RT).
- Perform resuspension of the cells.
- Discard the supernatant using a vacuum system (carefully, without disturbing the pellet) and resuspend the pellet in the transplantation medium containing 10 µM of RI, 1 µg/mL of DNase, and 2 µg/µL of DOX.
- Count the total cells in the suspension using a counting chamber and a manual cell counter and adjust the volume to a final concentration of 100,000 cells/µL.
- Keep the cell suspension in a closed tube on ice until transplantation for a maximum of 4 h.
NOTE: The transplantation has to be performed on the same day as the preparation of the cell suspension (7 DIV).
3. Intrahippocampal cell transplantation
NOTE: All the steps in this section are performed outside the cell culture hood in the animal facility. Early postnatal transplantation of cells into the brain was performed on P2, considering P0 the day of birth.
- Prepare for the transplantation experiment following the steps below.
- Autoclave all the surgical material or, if not possible, sterilize with another approved method.
- Mount the injecting syringe in the holder without any glass capillary. Rinse the needle with water.
NOTE: The needle must be 33 G. - Curve 90° the tip of a 30 G insulin needle.
- Create a homemade Play-Doh-like stage for positioning the pup with the head flat (Figure 2A).
- Fetch the mice cage with the mother and pups before preparing anything for the surgery to allow them to acclimatize to the new environment.
- Prepare a tray with wet ice and an empty cage with some paper for the isoflurane.
- Anesthetize the mouse pup.
- Soak a piece of paper with isoflurane (see Table of Materials) and place it inside the empty cage. Carefully take one pup and place it into the cage with the paper soaked in isoflurane.
NOTE: Never leave less than three pups with the mum. Do not exceed 20-30 s of isoflurane anesthesia, or the pup may die. - Verify the effect of the anesthesia by movement cessation.
- Immediately after anesthesia, place the pup onto a wet tissue on the surface of wet ice. Keep the animal on ice for 3 min until the upper limbs become whiteish (Figure 2B, blue arrow). This usually takes 2-3 min.
- Use this time for loading the syringe with the cell suspension. Resuspend the cells carefully with a pipette before.
- Place the pup on the stereotaxic frame. Use the ear bars in the opposite direction (Figure 2A, magenta arrows).
NOTE: The back side of the ear bars is less pointy, and as P2 pups do not have ears, use the flatter side to avoid hurting the animal. - From the side, check if the head is straight. The head must be flat; use the Play-Doh-like stage to adjust for that.
NOTE: Pups at this age have not yet opened their eyes, so no additional step in this regard needs to be done. Pups do not have fur at this age, so no shaving of the area is required. No specific pain treatment is given as it is not an open incision to the skin, and no suturing is involved. - Keep the pup covered on ice (on top of tissue paper) for the entire duration of the surgery.
NOTE: Do not place the ice in direct contact with the pup's skin.
- Soak a piece of paper with isoflurane (see Table of Materials) and place it inside the empty cage. Carefully take one pup and place it into the cage with the paper soaked in isoflurane.
- Perform the injection.
- Clean the surface of the skin using a soft tissue soaked in ethanol.
- Identify lambda and set the coordinates to zero on the digital display console of the stereotaxic instrument (Figure 2C, yellow dashed line). Sagittal and lambdoid sutures are easily visible by the eye, since they are vascularized, as red lines.
NOTE: If experiencing difficulties visualizing lambda, place a small piece of ice on the skin and wait a couple of minutes for it to cool down. Then, remove the piece of ice, and the subtle whitening of the skin will allow you to visualize. - Relocate the injection needle to the desired coordinates. In the described protocol, the targeted region was the hippocampus, and the coordinates were as follows: anterior-posterior (AP) +0.85 and medio-lateral (ML) +1.35.
- Use a 90° bent insulin needle to penetrate the skull and create a tiny hole.
- Bring down the injection needle until it has crossed the skull, and zero the dorso-ventral (DV) coordinates. Lower the needle until the desired DV coordinates. In the described protocol, the coordinates were DV −1.1.
- Inject according to the predetermined time intervals.
NOTE: In the described protocol, the exact timings were the following: (i) after going down to the DV coordinate, wait 3 min, (ii) inject 1 µL of volume for 5 min, and (iii) after all the volume has been injected, wait 3 min. - Retract the needle slowly.
- End the procedure.
NOTE: The surgery cannot last more than 15 min to ensure the pup's survival and avoid any damage derived from the anesthesia by hypothermia.- Warm up the pup with the hands until it starts moving before giving it back to the mother.
- Give DOX at a concentration of 1 mg/mL in 0.5% sucrose solution as drinking water for at least 2 days before and 3 weeks post transplantation (PT) to continue the cell differentiation in vivo.
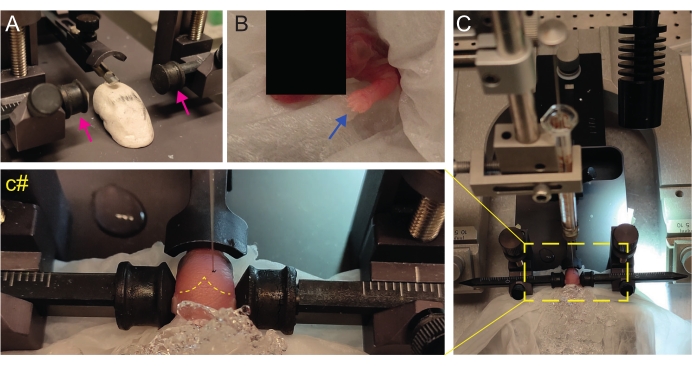
Figure 2: Stereotaxic transplantation in newborn mice pups at P2. (A) A Play-Doh-like stage for holding the pup's body in position and inverted ear bars (magenta arrows). (B) White front paw (blue arrow) indicative of the reduced blood flow in that area so that the pup is experiencing anesthesia by hypothermia. (C) Overview of the setup with the pup already covered by ice over the soft tissue paper. (c#) Closed-zoom of the head of the pup, with the injection needle already inserted into the brain (yellow dashed line indicating lambda and lambdoid sutures). This figure is adapted from Gonzalez Ramos et al.27. Please click here to view a larger version of this figure.
Following the protocol presented here and illustrated in Figure 1A, hdIN precursors were not proliferative yet at 7 DIV as defined by (i) negative immunoreactivity for the cell cycle marker Ki67 and (ii) expressing neuronal markers such as microtubule-associated protein 2 (MAP2) (Figure 1B–E). This characterization was performed on leftover cells replated for 24 h after having undergone all the procedure steps. In addition, the gene expression analysis published previously indicated that a rapid transition from the pluripotent state to a neuronal phenotype occurs around 4 DIV and 7 DIV8. Overall, these results confirmed the presence of postmitotic cells and the absence of risk for teratoma formation.
Next, the hdIN precursors' survival after early postnatal transplantation into the hippocampus of wild-type (WT) mice was tested by immunohistochemistry against the human cytoplasmatic marker STEM121. The hdIN precursors were transplanted into the right dorsal hippocampus of naïve immunocompetent mice at P2, which were then sacrificed at P14 and 2 months PT. Grafted cells were found across the whole dorsal hippocampus, as well as dispersed through the corpus callosum and the contralateral hippocampus, at both time points. Moreover, at both time points, grafted hdINs expressed Ascl1, one of the induction transcription factors (Supplementary Figure 1), and were not proliferative, as indicated by the absence of Ki67 expression (Supplementary Figure 2).
Importantly, no immune reaction or local inflammation against the transplanted cells was found either at P14 or 2 months PT, as assessed by the absence of reactive microglia identified using Iba1, CD68, and galectin-3 (Gal3) (Figure 3), the extent of astrogliosis determined by the glial fibrillary acidic protein (GFAP) and inflammatory cytokines such as interleukine-1 (IL-1), and the absence of cytotoxic T lymphocytes (CD8) (Figure 4).
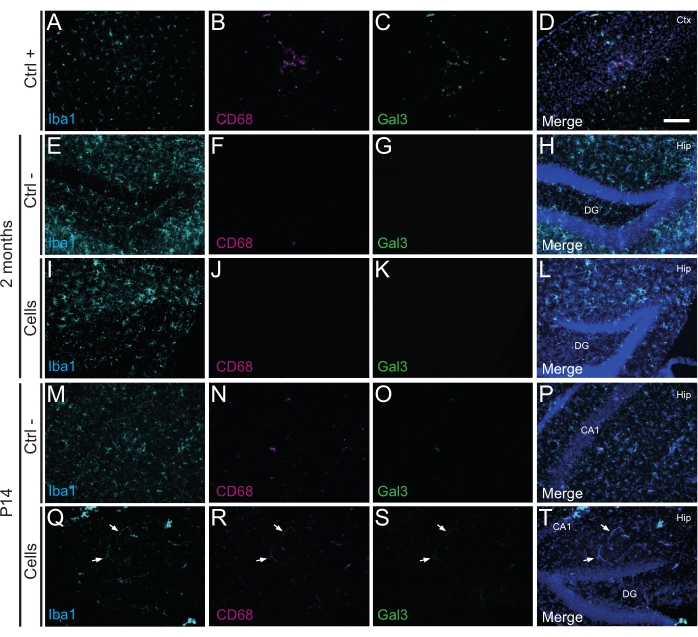
Figure 3: hdINs at P14 and 2 months PT into the hippocampus of newborn WT mice without triggering immune rejection from the host tissue. Immunofluorescence for Iba1, CD68, and Gal3 markers in brain tissue from (A–D) the proximity of an ischemic core area in an electrocoagulation stroke mouse model (positive control, Ctrl +), (E–H) negative control animals (Ctrl-) at 2 months and (M–P) P14, and (I–L) animals that have undergone cell transplantation at 2 months and (Q–T) P14. The white arrows indicate some examples of blood vessels visible at all channels due to autofluorescence. Abbreviations: Ctx = cortex; Hip = hippocampus; DG = dentate gyrus; CA1 = cornu ammonis 1. Scale bar: 50 µm. Please click here to view a larger version of this figure.
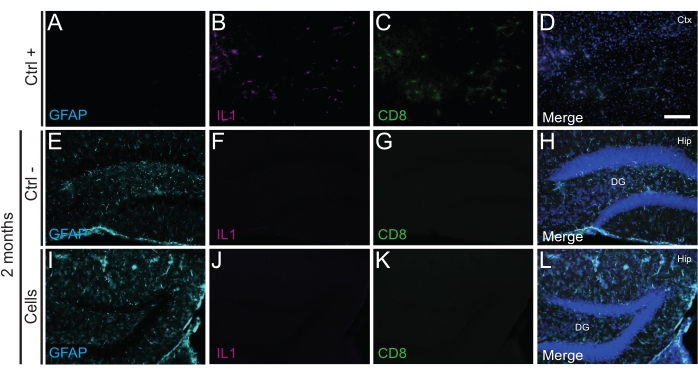
Figure 4: hdINs at 2 months PT into the hippocampus of newborn WT mice without triggering immune rejection from the host tissue. Immunofluorescence for IL1, GFAP, and CD8 markers in brain tissue from (A–D) the proximity of an ischemic core area in an electrocoagulation stroke mouse model (positive control, Ctrl +), (E–H) negative control animals (Ctrl-) at 2 months, and (I-L)animals that have undergone cell transplantation at 2 months. Abbreviations: Ctx = cortex; Hip = hippocampus; DG = dentate gyrus. Scale bar: 50 µm. Please click here to view a larger version of this figure.
Similarly, hdIN precursors were also transplanted into the hippocampus of Cntnap2 KO mice, a model for autism spectrum disorder and cortical dysplasia-focal epilepsy syndrome. In the Cntnap2 KO mice, indeed, the hdINs survived up to 9 months PT and were localized at the injection site, although they were also dispersed across the ipsilateral and even the contralateral hippocampus as observed in the WT mice (Figure 5). Moreover, most of the grafted hdINs were immunoreactive for interneuron markers, as expected from previous results in vitro8,26 and in adult rodents in vivo25.
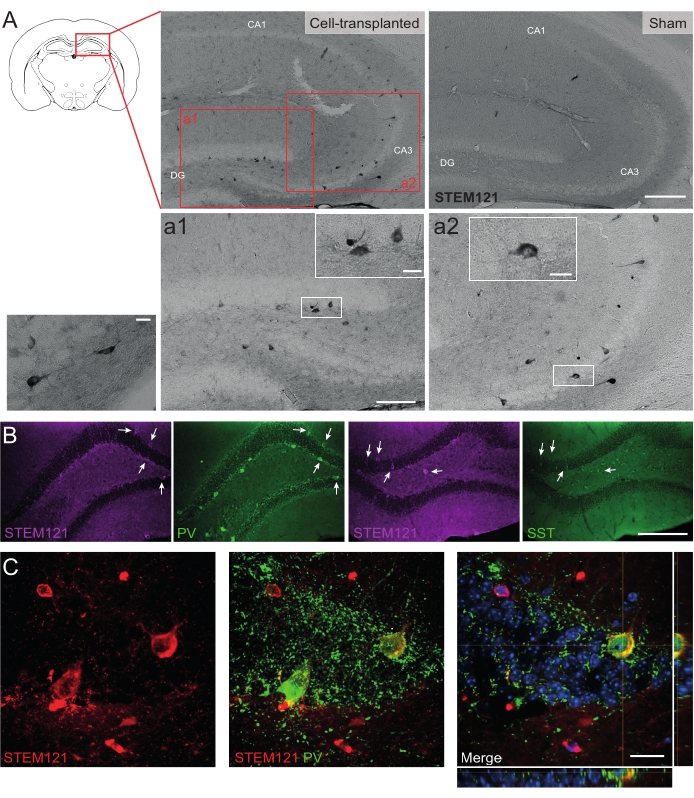
Figure 5: Grafted hdINs in the hippocampus of Cntnap2 KO mice at 9 months PT. (A) Immunochemistry against the cytoplasmatic human marker STEM121 in cell-transplanted (left) and sham (right) mice. (a1 and a2) Magnified images of STEM121 positive cells. (B) Immunofluorescence for STEM121 (magenta) and the interneuron markers parvalbumin (PV) and somatostatin (SST). White arrows indicate double positive cells for STEM121 and the respective interneuron marker. (C) Orthogonal view of a grafted hdIN immunoreactive for STEM121 and PV. Scale bar: 200 µm (A and B), 100 µm (a1 and a2), 20 µm (small square magnification in a1 and a2, and C). Please click here to view a larger version of this figure.
Supplementary Figure 1: Grafted hdINs at 2 months PT into the hippocampus of newborn WT mice expressing Ascl1. Immunofluorescence against Ascl1 and the cytoplasmatic human marker STEM121 at the (A) CA3 and (B) DG in cell-transplanted WT mice. (a) Magnified image of a STEM121 positive cell. The white arrows indicate double-positive cells for STEM121 and Ascl1. Scale bar: 100 µm. Please click here to download this File.
Supplementary Figure 2: Post-mitotic grafted hdINs at 2 months PT into the hippocampus of newborn WT mice. Immunofluorescence against the proliferative marker Ki67 and the cytoplasmatic human marker STEM121 in (A) naïve and (B) cell-transplanted WT mice. (b) Magnified image of a STEM121 positive cell. The yellow arrow indicates a cell positive for Ki67 and negative for STEM121. The white arrow indicates cells positive for STEM121 and negative for Ki67. The white asterisk points out a lateral ventricle. Scale bar: 100 µm. Please click here to download this File.
