番茄(番茄 )是世界上最重要的蔬菜作物之一,2020年产量为1.868亿吨肉质水果,面积达510万公顷1。它属于大型茄科,约有2,716种2,包括许多具有重要商业意义的作物,如茄子,辣椒,马铃薯和烟草。栽培番茄是一种二倍体物种(2n = 2x = 24),基因组大小约为900 Mb3。长期以来,通过从野生茄属中选择理想的性状,在番 茄 驯化和育种方面做出了巨大努力。番茄遗传学资源中心列出了 5,000 多种番茄种质,全球储存了 80,000 多种番茄种质4.番茄植株在温室中多年生,通过种子繁殖。成熟的番茄种子由三个主要隔室组成:一个成熟的胚胎、残留的细胞型胚乳和一个坚硬的种皮5,6 (图 1A)。双重受精后,细胞型胚乳的发育先于受精卵的发育。在开花后~5-6天(DAF),当胚乳由6至8个细胞核7组成时,首先观察到双细胞前胚。在 茄子松中,胚胎在20 DAF后接近其最终大小,种子在32 DAF8后可以发芽。随着胚胎的发育,胚乳逐渐被吸收,种子中只剩下少量胚乳。残留胚乳由胚根尖端周围的小胚乳和种子其余部分的侧胚乳组成9,10。外种皮由外皮增厚木质化的外表皮发育而来,与外皮残余物的死层形成硬壳以保护胚胎和胚乳5。
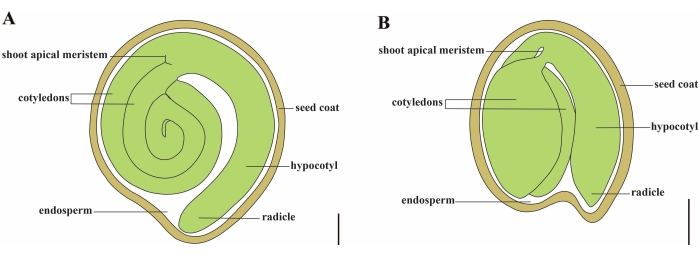
图1: Solanum lycopersicum 和 拟南芥中 成熟种子的示意图。 (A)成熟番茄种子的纵向解剖结构。(B)成熟 拟南芥 种子的纵向解剖。成熟的番茄种子的大小大约是 拟南芥 种子的 70 倍。比例尺 = (A) 400 μm, (B) 100 μm。 请点击此处查看此图的大图。
高质量番茄种子的生产取决于胚胎、胚乳和母体种子成分之间的协调11.剖析种子发育中的关键基因和网络需要对突变种子进行深入和完整的表型记录。传统的包埋切片技术,如半薄切片和石蜡切片,被广泛用于番茄种子,以观察胚胎的局部和更精细的结构12,13,14,15。然而,从薄片分析种子发育通常很费力,并且缺乏z轴空间分辨率。相比之下,组织清除是一种快速有效的方法,可以查明最有可能发生的胚胎缺陷的发育阶段16。清除方法通过用一种或多种生化剂均质化折射率来降低内部组织的不透明度16。全组织清除可以在不破坏其完整性的情况下观察植物组织结构,并且清除技术和三维成像的结合已成为获取植物器官形态和发育状态信息的理想解决方案17,18。多年来,种子清除技术已用于各种植物物种,包括拟南芥、大麦和普通贝塔19、20、21、22、23。其中,全镶样胚珠清除技术因其体积小、种皮细胞4-5层、核型胚乳24、25等特点,成为研究拟南芥种子发育的有效方法。随着不同清除混合物的不断更新,例如Hoyer溶液26的出现,尽管大麦胚珠的胚乳占种子的大部分,但大麦胚珠的内部结构以高度清晰的方式成像。甜菜的胚胎发生可以通过清除结合真空处理和盐酸软化来观察19。尽管如此,与上述物种不同,尚未报道通过清除番茄种子协议进行的胚胎学观察。这阻碍了对西红柿胚胎和种子发育的详细调查。
水合氯醛通常用作澄清溶液,其允许浸没的组织和细胞在不同的光学平面上显示,并基本上保留细胞或组织成分27,28,29。基于水合氯醛的清除方案已成功用于种子的全安装清除,以观察拟南芥21,28的胚胎和胚乳。然而,这种清除溶液在清除番茄种子方面效率不高,番茄种子比拟南芥种子更不透水。物理屏障包括:(1)番茄外皮在3至15 DAF 30,31处有近20个细胞层,(2)番茄胚乳是细胞型,而不是核型32,(3)番茄种子的大小约为70倍33,34和(4)产生大量的种皮粘液,这阻止了清除试剂的渗透并影响了胚胎细胞的可视化。
因此,本报告提出了一种优化的基于水合氯醛的清除方法,用于在不同阶段对番茄种子进行整体安装清除,该方法可以对胚胎发育过程进行深度成像(图2)。
