Tomat (S. lycopersicum L.) er en af de vigtigste vegetabilske afgrøder rundt om i verden med en produktion på 186,8 millioner tons kødfulde frugter fra 5,1 millioner hektar i 20201. Det tilhører den store Solanaceae-familie med omkring 2.716 arter2, herunder mange kommercielt vigtige afgrøder som aubergine, peberfrugter, kartoffel og tobak. Den dyrkede tomat er en diploid art (2n = 2x = 24) med en genomstørrelse på ca. 900 Mb3. I lang tid er der gjort en stor indsats for tomat domesticering og avl ved at vælge ønskelige træk fra vilde Solanum spp. Der er over 5.000 tomattiltrædelser opført i Tomato Genetics Resource Center, og mere end 80.000 kimplasmer af tomater opbevares over hele verden4. Tomatplanten er flerårig i drivhuset og formeres af frø. Et modent tomatfrø består af tre hovedrum: et fuldvoksent embryo, endosperm af cellulær type og en hård frøbelægning 5,6 (figur 1A). Efter dobbelt befrugtning går udviklingen af endosperm af cellulær type forud for udviklingen af zygoter. Ved ~ 5-6 dage efter blomstring (DAF) observeres tocellet proembryo først, når endospermen består af seks til otte kerner7. I Solanum pimpinellifolium nærmer embryoet sin endelige størrelse efter 20 DAF, og frø er levedygtige til spiring efter 32 DAF8. Efterhånden som embryoet udvikler sig, absorberes endospermen gradvist, og kun en lille mængde endosperm forbliver i frøet. Den resterende endosperm består af mikropylar endosperm omkring radikelspidsen og lateral endosperm i resten af frøet 9,10. Den ydre frøbeklædning er udviklet fra fortykket og lignificeret ydre epidermis af integumentet, og med de døde lag af integumentrester danner de en hård skal for at beskytte embryoet og endospermen5.
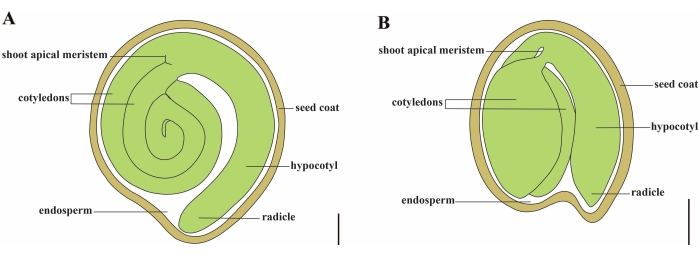
Figur 1: Skematisk repræsentation af et modent frø i Solanum lycopersicum og Arabidopsis thaliana. (A) Langsgående anatomi af et modent tomatfrø. (B) Langsgående anatomi af et modent Arabidopsisfrø. Et modent tomatfrø er ca. 70 gange større end et Arabidopsisfrø. Skalabjælker = (A) 400 μm, (B) 100 μm. Klik her for at se en større version af denne figur.
Produktion af tomatfrø af høj kvalitet afhænger af koordineringen mellem embryoet, endospermen og moderens frøkomponenter11. Dissekering af nøglegener og netværk i frøudvikling kræver en dyb og fuldsporet fænotypisk registrering af mutante frø. Konventionelle indlejrings-sektioneringsteknikker, såsom den halvtynde sektion og paraffinsektionen, anvendes bredt på tomatfrø for at observere de lokale og finere strukturer i embryoet12,13,14,15. Imidlertid er analyse af frøudviklingen fra tynde sektioner normalt besværlig og mangler z-akse rumlig opløsning. Til sammenligning er vævsrensning en hurtig og effektiv metode til at lokalisere udviklingsstadiet af embryodefekter, der mest sandsynligt vil forekomme16. Rydningsmetoden reducerer uigennemsigtigheden af internt væv ved at homogenisere brydningsindekset med et eller flere biokemiske midler16. Helvævsrensning tillader observation af en plantevævsstruktur uden at ødelægge dens integritet, og kombinationen af clearingteknologi og tredimensionel billeddannelse er blevet en ideel løsning til at få information om morfologien og udviklingstilstanden af et planteorgan17,18. I årenes løb er frørensningsteknikker blevet brugt i forskellige plantearter, herunder Arabidopsis thaliana, Hordeum vulgare og Beta vulgaris 19,20,21,22,23. Blandt disse har helmonteret ægløsningsteknologi været en effektiv tilgang til at studere frøudvikling af Arabidopsis på grund af dens lille størrelse, 4-5 lag af frøbeklædningscellen og endospermen af nuklear type24,25. Med den løbende opdatering af forskellige rydningsblandinger, såsom fremkomsten af Hoyers opløsning26, blev de indre strukturer af bygæglet afbildet med en høj grad af klarhed, selvom dets endosperm udgør størstedelen af frøene. Embryogenese af sukkerroer kan observeres ved rydning kombineret med vakuumbehandling og blødgøring med saltsyre19. I modsætning til ovennævnte arter er der imidlertid ikke rapporteret om embryologiske observationer ved rydning af protokoller i tomatfrø. Dette forhindrer detaljeret undersøgelse af tomaternes embryonale og frøudvikling.
Klorhydrat anvendes almindeligvis som en rydningsopløsning, der gør det muligt at vise de nedsænkede væv og celler på forskellige optiske planer og bevarer i væsentlig grad cellerne eller vævskomponenterne27,28,29. Klorhydratbaseret clearingprotokol er med succes blevet brugt til helmontering af frø for at observere embryoet og endospermen af Arabidopsis21,28. Denne rydningsløsning er imidlertid ikke effektiv til at rydde tomatfrø, som er mere uigennemtrængelige end Arabidopsisfrø. De fysiske barrierer omfatter: (1) tomatintegumentet har næsten 20 cellelag ved 3 til 15 DAF 30,31, (2) tomatendospermen er cellulær type, ikke nuklear type32, og (3) tomatfrø er ca. 70 gange større i størrelse33,34 og (4) producerer store mængder frøbelægningsslimhinde, som blokerer penetrationen af rydningsreagenser og påvirker visualiseringen af embryoceller.
Derfor præsenterer denne rapport en optimeret klorhydratbaseret rydningsmetode til helmonteringsrensning af tomatfrø på forskellige stadier, hvilket muliggør dyb billeddannelse af embryoudviklingsprocessen (figur 2).
