Tomaat (S. lycopersicum L.) is een van de belangrijkste groentegewassen ter wereld, met een output van 186,8 miljoen ton vlezig fruit van 5,1 miljoen hectare in 20201. Het behoort tot de grote Solanaceae-familie met ongeveer 2.716 soorten2, waaronder veel commercieel belangrijke gewassen zoals aubergine, paprika’s, aardappelen en tabak. De gekweekte tomaat is een diploïde soort (2n = 2x = 24) met een genoomgrootte van ongeveer 900 Mb3. Lange tijd is er veel moeite gedaan voor het domesticeren en veredelen van tomaten door het selecteren van gewenste eigenschappen uit wilde Solanum spp. Er zijn meer dan 5.000 tomatentoetredingen vermeld in het Tomato Genetics Resource Center en meer dan 80.000 kiemplasma van tomaten worden wereldwijd opgeslagen4. De tomatenplant is overblijvend in de kas en vermeerdert zich door zaden. Een rijp tomatenzaad bestaat uit drie belangrijke compartimenten: een volgroeid embryo, resterend cellulair endosperm en een harde zaadlaag 5,6 (figuur 1A). Na dubbele bevruchting gaat de ontwikkeling van cellulair endosperm vooraf aan de ontwikkeling van zygoten. Bij ~5-6 dagen na de bloei (DAF) wordt tweecellige proembryo voor het eerst waargenomen wanneer het endosperm bestaat uit zes tot acht kernen7. In Solanum pimpinellifolium nadert het embryo zijn uiteindelijke grootte na 20 DAF en zijn zaden levensvatbaar voor ontkieming na 32 DAF8. Naarmate het embryo zich ontwikkelt, wordt het endosperm geleidelijk geabsorbeerd en blijft er slechts een kleine hoeveelheid endosperm in het zaad achter. Het resterende endosperm bestaat uit micropylar endosperm rond de radikelpunt en lateraal endosperm in de rest van het zaad 9,10. De buitenste zaadlaag is ontwikkeld uit verdikte en gelamineerde buitenste epidermis van het integument, en met de dode lagen van integumentresten vormen ze een harde schaal om het embryo en endosperm5 te beschermen.
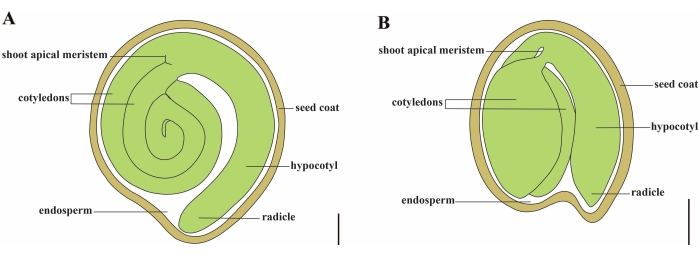
Figuur 1: Schematische weergave van een rijp zaad in Solanum lycopersicum en Arabidopsis thaliana. (A) Longitudinale anatomie van een rijp tomatenzaad. (B) Longitudinale anatomie van een volwassen Arabidopsis zaad. Een volwassen tomatenzaad is ongeveer 70 keer groter dan een Arabidopsiszaadje . Schaalbalken = (A) 400 μm, (B) 100 μm. Klik hier om een grotere versie van dit cijfer te bekijken.
De productie van hoogwaardige tomatenzaden hangt af van de coördinatie tussen het embryo, het endosperm en de maternale zaadcomponenten11. Het ontleden van belangrijke genen en netwerken in zaadontwikkeling vereist een diepe en full-track fenotypische opname van gemuteerde zaden. Conventionele inbeddingstechnieken, zoals de halfdunne sectie en paraffinesectie, worden op grote schaal toegepast op tomatenzaden om de lokale en fijnere structuren van het embryo te observeren 12,13,14,15. Het analyseren van de zaadontwikkeling vanuit dunne secties is echter meestal bewerkelijk en mist de ruimtelijke resolutie van de z-as. Ter vergelijking: weefselopruiming is een snelle en efficiënte methode om het ontwikkelingsstadium van embryodefecten te lokaliseren die het meest waarschijnlijk optreden16. De clearingmethode vermindert de ondoorzichtigheid van inwendig weefsel door de brekingsindex te homogeniseren met een of meer biochemische agentia16. Het opruimen van hele weefsels maakt observatie van een plantenweefselstructuur mogelijk zonder de integriteit ervan te vernietigen, en de combinatie van clearingtechnologie en driedimensionale beeldvorming is een ideale oplossing geworden om informatie te verkrijgen over de morfologie en ontwikkelingstoestand van een plantenorgaan17,18. In de loop der jaren zijn zaadzuiveringstechnieken gebruikt bij verschillende plantensoorten, waaronder Arabidopsis thaliana, Hordeum vulgare en Beta vulgaris 19,20,21,22,23. Onder deze, de hele-mount ovule clearing technologie is een efficiënte benadering geweest voor het bestuderen van zaadontwikkeling van Arabidopsis, vanwege zijn kleine formaat, 4-5 lagen van de zaadlaagcel, en het nucleaire type endosperm24,25. Met de voortdurende actualisering van verschillende clearingmengsels, zoals de opkomst van Hoyer’s oplossing26, werden interne structuren van de gerst-eicel met een hoge mate van helderheid in beeld gebracht, hoewel het endosperm het grootste deel van de zaden uitmaakt. Embryogenese van suikerbieten kan worden waargenomen door opklaring in combinatie met vacuümbehandeling en verzachting met zoutzuur19. Niettemin zijn, in tegenstelling tot de hierboven genoemde soorten, embryologische waarnemingen door clearingprotocollen in tomatenzaden niet gemeld. Dit voorkomt gedetailleerd onderzoek naar de embryonale en zaadontwikkeling van tomaten.
Chloraalhydraat wordt vaak gebruikt als een opruimoplossing waarmee de ondergedompelde weefsels en cellen op verschillende optische vlakken kunnen worden weergegeven en de cellen of weefselcomponenten aanzienlijk behouden 27,28,29. Het op chloraalhydraat gebaseerde clearingprotocol is met succes gebruikt voor het volledig opruimen van zaden om het embryo en endosperm van Arabidopsis21,28 te observeren. Deze clearingoplossing is echter niet efficiënt bij het opruimen van tomatenzaden, die ondoordringbaarder zijn dan Arabidopsis-zaden. De fysieke barrières omvatten: (1) het tomatenintegument heeft bijna 20 cellagen bij 3 tot 15 DAF30,31, (2) het tomatenendosperm is cellulair type, niet nucleair type32, en (3) tomatenzaden zijn ongeveer 70 keer groter in grootte33,34 en (4) produceren grote hoeveelheden zaadlaagslijm, wat de penetratie van clearingreagentia blokkeert en de visualisatie van embryocellen beïnvloedt.
Daarom presenteert dit rapport een geoptimaliseerde op chloraalhydraat gebaseerde clearingmethode voor het volledig opruimen van tomatenzaden in verschillende stadia, waardoor diepe beeldvorming van het embryo-ontwikkelingsproces mogelijk is (figuur 2).
