Tomat (S. lycopersicum L.) är en av de viktigaste grönsaksgrödorna runt om i världen, med en produktion på 186,8 miljoner ton köttiga frukter från 5,1 miljoner hektar 20201. Den tillhör den stora familjen Solanaceae med cirka 2 716 arter2, inklusive många kommersiellt viktiga grödor som aubergine, paprika, potatis och tobak. Den odlade tomaten är en diploid art (2n = 2x = 24) med en genomstorlek på cirka 900 Mb3. Under lång tid har stora ansträngningar gjorts för tomattämjning och avel genom att välja önskvärda egenskaper från vilda Solanum spp. Det finns över 5 000 tomatanslutningar listade i Tomato Genetics Resource Center och mer än 80 000 bakterieplasma av tomater lagras över hela världen4. Tomatplantan är flerårig i växthuset och förökas av frön. Ett moget tomatfrö består av tre huvudfack: ett fullvuxet embryo, kvarvarande endosperm av cellulär typ och en hårdfröbeläggning 5,6 (figur 1A). Efter dubbel befruktning föregår utvecklingen av endosperm av cellulär typ utvecklingen av zygoter. Vid ~ 5-6 dagar efter blomning (DAF) observeras tvåcellig proembryo först när endospermen består av sex till åtta kärnor7. I Solanum pimpinellifolium närmar sig embryot sin slutliga storlek efter 20 DAF, och frön är livskraftiga för spiring efter 32 DAF8. När embryot utvecklas absorberas endospermen gradvis och endast en liten mängd endosperm kvarstår i fröet. Den återstående endospermen består av mikropylar endosperm som omger radikelspetsen och lateral endosperm i resten av fröet 9,10. Den yttre fröbeläggningen är utvecklad från förtjockad och lignifierad yttre epidermis av integumentet, och med de döda lagren av integumentrester bildar de ett hårt skal för att skydda embryot och endospermen5.
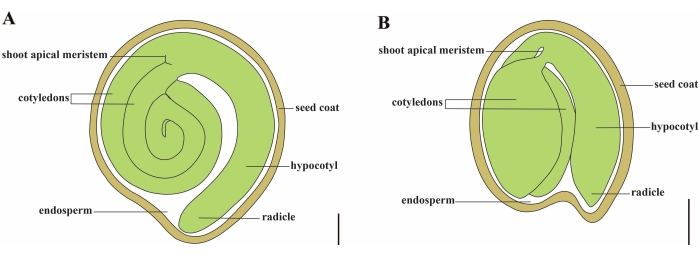
Figur 1: Schematisk representation av ett moget frö i Solanum lycopersicum och Arabidopsis thaliana. (A) Longitudinell anatomi hos ett moget tomatfrö. B) Longitudinell anatomi hos ett moget Arabidopsisfrö. Ett moget tomatfrö är ungefär 70 gånger större än ett Arabidopsis-frö. Skalstänger = (A) 400 μm, (B) 100 μm. Klicka här för att se en större version av denna figur.
Produktion av högkvalitativa tomatfrön beror på samordningen mellan embryot, endospermen och moderfrökomponenterna11. Att dissekera viktiga gener och nätverk i fröutveckling kräver en djup och fullständig fenotypisk registrering av mutanta frön. Konventionella inbäddnings-sektionstekniker, såsom den halvtunna sektionen och paraffinsektionen, används i stor utsträckning på tomatfrön för att observera de lokala och finare strukturerna hos embryot12,13,14,15. Att analysera fröutvecklingen från tunna sektioner är dock vanligtvis mödosamt och saknar z-axelns rumsliga upplösning. I jämförelse är vävnadsrensning en snabb och effektiv metod för att fastställa utvecklingsstadiet för embryodefekter som mest sannolikt kommer att inträffa16. Clearingmetoden minskar ogenomskinligheten hos inre vävnad genom att homogenisera brytningsindexet med ett eller flera biokemiska medel16. Rensning av hela vävnader möjliggör observation av en växtvävnadsstruktur utan att förstöra dess integritet, och kombinationen av röjningsteknik och tredimensionell avbildning har blivit en idealisk lösning för att få information om morfologin och utvecklingstillståndet hos ett växtorgan17,18. Under åren har fröröjningstekniker använts i olika växtarter, inklusive Arabidopsis thaliana, Hordeum vulgare och Beta vulgaris 19,20,21,22,23. Bland dessa har den helmonterade ägglossningstekniken varit ett effektivt tillvägagångssätt för att studera fröutveckling av Arabidopsis, på grund av dess lilla storlek, 4-5 lager av fröskiktcellen och endospermen av kärntyp24,25. Med den kontinuerliga uppdateringen av olika clearingblandningar, såsom framväxten av Hoyers lösning26, avbildades inre strukturer i kornäggstocken med hög grad av klarhet även om dess endosperm utgör huvuddelen av fröna. Embryogenes av sockerbetor kan observeras genom rensning i kombination med vakuumbehandling och mjukning med saltsyra19. Till skillnad från de arter som nämns ovan har dock embryologiska observationer genom rensningsprotokoll i tomatfrön inte rapporterats. Detta förhindrar detaljerad undersökning av embryonal och fröutveckling av tomater.
Klorhydrat används ofta som en clearinglösning som gör att de nedsänkta vävnaderna och cellerna kan visas på olika optiska plan och bevarar cellerna eller vävnadskomponenterna27,28,29. Klorhydratbaserat clearingprotokoll har framgångsrikt använts för helmontering av frön för att observera embryot och endospermen av Arabidopsis21,28. Denna clearinglösning är emellertid inte effektiv för att rensa tomatfrön, som är mer ogenomträngliga än Arabidopsisfrön. De fysiska barriärerna inkluderar: (1) tomatintegumentet har nästan 20 cellskikt vid 3 till 15 DAF 30,31, (2) tomatendospermen är cellulär, inte kärntyp32, och (3) tomatfrön är cirka 70 gånger större i storlek33,34 och (4) producerar stora mängder fröskiktsmucilage, vilket blockerar penetrationen av clearingreagens och påverkar visualiseringen av embryoceller.
Därför presenterar denna rapport en optimerad klorhydratbaserad röjningsmetod för helmonterad rensning av tomatfrön i olika stadier, vilket möjliggör djup avbildning av embryoutvecklingsprocessen (figur 2).
