AdipoSPH mice were developed by breeding SPH and Adipoq-Cre mouse strains. Both mouse strains were in a hybrid C57BL6J-DBA/2J background (according to the commercial supplier; see Table of Materials). The SPH mouse lineage was originally described by Zhou et al.14.
In vivo beige adipocyte development through AdipoSPH-mediated Prdm16 overexpression
To evaluate the capacity of the model described in this study to develop beige adipocytes in vivo, the AAV carrying the sgRNA targeting Prdm16 gene was injected into the iWAT of AdipoSPH mice. Prdm16 is a well-established transcription factor that determines beige adipocyte development and function20,21. It is important to mention that here we chose a previously tested and validated sgRNA targeting the endogenous Prdm16 gene14. AAV carrying an empty sgRNA was used as a control. At 10 days after AAV injection, iWAT was harvested for histological and gene expression analyses (Figure 1A). As expected, immunofluorescence images of iWAT from AdipoSPH mice demonstrated the expression of dCas9 in both the control and Prdm16 groups (Figure 1B). Additionally, mCherry expression confirmed the success of AAV infection of iWAT in both the control and Prdm16 groups (Figure 1B). SPH-induced Prdm16 expression clearly induced a widespread accumulation of multilocular beige adipocytes into iWAT (Figure 1B). Moreover, quantitative PCR (qPCR) revealed an increased expression of Prdm16 and the thermogenic gene program (Ucp1, Cox8b, Ppargc1a, and Cidea) in the Prdm16 group compared to the control (Figure 1C).
Promotion of beige adipocyte development and enhancement of oxygen consumption in vitro by AdipoSPH-mediated Prdm16 overexpression
Next, the suitability of this model for studying beige adipocyte biology in vitro was investigated. To this end, preadipocytes derived from the iWAT of AdipoSPH mice were transduced with AAV carrying the sgRNA sequence targeting Prdm16. At seven days after differentiation, beige adipocytes were used for the analysis of gene expression and oxygen consumption (Figure 2A). Empty sgRNA was used as a control. It is worth noting that the removal of STOP codon by CRE recombinase and activation of SPH machinery was under the control of the adiponectin promoter/enhancer, resulting in the activation of the endogenous Prdm16 gene in mature adipocytes. Gene expression analysis confirmed the increased expression of the endogenous Prdm16 gene and thermogenic genes in the Prdm16 overexpression group compared to the control (Figure 2B). To confirm that the SPH-induced beige adipocytes were functionally thermogenic, a high-resolution respirometry assay was performed on the primary adipocytes. The respirometry data analysis and interpretation was performed as recently described22. SPH-induced Prdm16 expression resulted in higher basal and maximal oxygen consumption than that in control cells (Figure 2C). Importantly, the data indicated enhanced uncoupled respiration (oligomycin-insensitive) in the Prdm16 group compared with that in control group (Figure 2C, Table 3). Taken together, these results revealed that SPH-induced expression of endogenous Prdm16 recapitulates the browning phenotype and enhances oxygen consumption rates observed in conventional Prdm16 Tg mice.
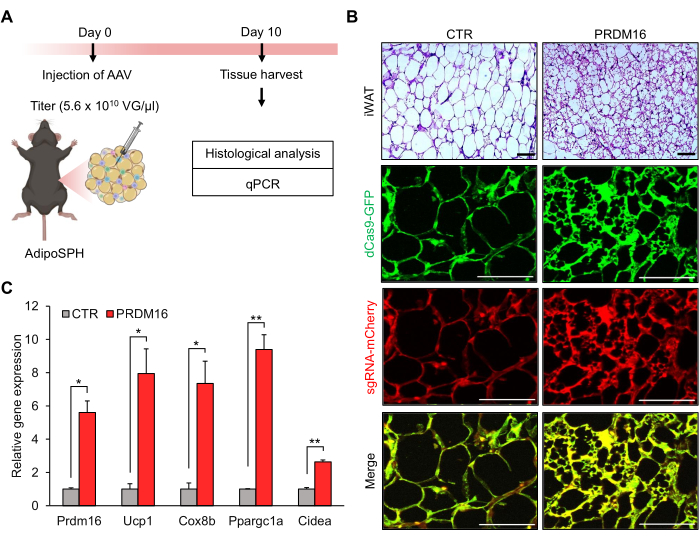
Figure 1: Beige fat development by injection of adeno-associated virus (AAV) targeting endogenous Prdm16 gene into inguinal white adipose tissue (iWAT) of AdipoSPH mice. (A) Schematic illustration of the in vivo beige adipocyte experimental design (created using Biorender.com). (B) Histological (hematoxylin and eosin [H&E] staining) images of iWAT. Scale bar = 50 µm. (C) Quantitative PCR (qPCR) of Prdm16 and thermogenic genes (Ucp1, Cox8b, Ppargc1α, and Cidea; n = 3). Data are presented as mean ± SEM. *p < 0.05; **p < 0.01 for Prdm16 versus control (CTR) by unpaired Student's t-test. Please click here to view a larger version of this figure.
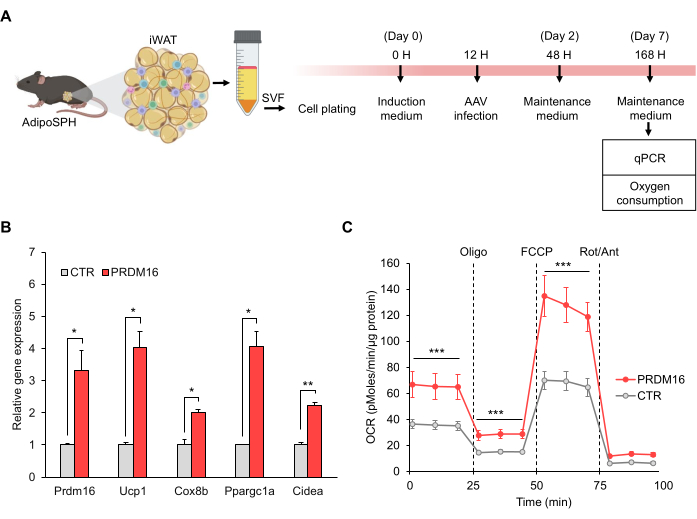
Figure 2: SPH-induced beige adipocyte differentiation by Prdm16 overexpression. (A) Schematic illustration of the in vitro beige adipocyte experimental design (created using Biorender.com). (B) Relative gene expression of Prdm16 and thermogenic genes (Ucp1, Cox8b, Ppargc1α, and Cidea; n = 3). (C) Oxygen consumption rate (OCR), n = 6. Data are presented as mean ± SEM. *p < 0.05; **p < 0.01; ***p < 0.001 for Prdm16 versus control (CTR) by (B) unpaired Student's t-test and (C) two-way repeated-measures ANOVA, followed by Tukey's test. Please click here to view a larger version of this figure.
Supplemental Figure 1: Sanger sequencing of Prdm16 sgRNA. (A) Schematic illustration demonstrating the alignment of single-stranded complementary Prdm16 oligonucleotides. (B) Sanger sequencing of Prdm16 sgRNA using universal primer. Please click here to download this File.
| Gene | Species | Forward | Reverse | |||||
| Gene expression | Prdm16 | mouse | CAGCACGGTGAAGCCATTC | GCGTGCATCCGCTTGTG | ||||
| Ucp1 | mouse | TCTCAGCCGGCTTAATGACTG | GGCTTGCATTCTGACCTTCAC | |||||
| Ppargc1a | mouse | AGCCGTGACCACTGACAACGAG | GCTGCATGGTTCTGAGTGCTAAG | |||||
| Cox8b | mouse | GAACCATGAAGCCAACGACT | GCGAAGTTCACAGTGGTTCC | |||||
| Cidea | mouse | ATCACAACTGGCCTGGTTACG | TACTACCCGGTGTCCATTTCT | |||||
| 36B4 | mouse | TCCAGGCTTTGGGCATCA | CTTTATCAGCTGCACATCACTCAGA | |||||
| Molecular cloning | sgRNA Prdm16 sequence | mouse | CGAGCTGCGCTGAAAAGGGG | CCCCTTTTCAGCGCAGCTCG | ||||
| universal primer | mouse | GAGGGCCTATTTCCC ATGATTCCTTCATAT |
||||||
Table 1: Primer sequences used in the study.
| Medium | Composition | |||
| Complete medium | Dulbecco's Modified Eagle Medium (DMEM) with L-Glutamine | |||
| 10% of fetal bovine serum (FBS) | ||||
| 2.5% of penicillin/streptomycin | ||||
| Induction medium | Complete medium | |||
| Indomethacin, final concentration 125 μM (0.125 M stock in ethanol). NOTE: Indomethacin must be heated to 90 °C for 10 seconds to be dissolved. | ||||
| Insulin, final concentration 20 nM (1 mM stock, Add 1 μL of HCl to solubilize (5.73 mg/mL). Store stock at -20 °C. | ||||
| Dexamethasone, final concentration 2 μg/mL (2 mg/mL stock in ethanol) | ||||
| 3-Isobutyl-1-methylxanthine (IBMX), final concentration 500 μM (0.25 M stock in 0.5 M KOH ) | ||||
| 3,3',5-Triiodo-L-thyronine (T3), final concentration 1 nM (10 μM stock, dissolve T3 in 1 M HCl and EtOH 1:4 to stock). | ||||
| Rosiglitazone, final concentration 1 μg/mL (1 mg/mL stock in ethanol) | ||||
| Maintenance medium | Complete medium | |||
| Insulin, final concentration 20 nM (1 mM stock). Add 1 μL of HCl to solubilize (5.73 mg/mL). | ||||
| Rosiglitazone, final concentration 1 μg/mL (1 mg/mL stock in ethanol) | ||||
Table 2: Composition of media used in the study.
| Parameters | CTR | PRDM16 | p-value | ||
| No mitochondrial oxygen consumption (pMoles/min/µg protein) | 8.19 ± 1.40 | 10.80 ± 1.83 | 0.01 | ||
| Basal respiration (pMoles/min/µg protein) | 28.82 ± 5.20 | 52.58 ± 13.73 | 0.001 | ||
| Maximal respiration (pMoles/min/µg protein) | 63.81 ± 9.80 | 122.94 ± 22.31 | < 0.001 | ||
| Spare respiratory capacity (pMoles/min/µg protein) | 34.98 ± 11.09 | 70.36 ± 26.06 | 0.006 | ||
| Oligo insensitive (pMoles/min/µg protein) | 8.27 ± 2.29 | 15.85 ± 5.48 | 0.005 | ||
| Oligo sensitive (pMoles/min/µg protein) | 20.54 ± 5.68 | 36.72 ± 14.79 | 0.016 | ||
| Coupling efficiency | 71.28 ± 1.51 | 69.84 ± 3.05 | 0.163 | ||
| Cell RCR | 7.70 ± 1.46 | 7.75 ± 2.43 | 0.485 | ||
Table 3: Respiratory parameters of SPH-induced beige adipocytes.
