التحويلات اليومية وأرشفة السكان وقياس اللياقة البدنية في تجربة التطور طويلة المدى مع الإشريكية القولونية
Summary
يصف هذا البروتوكول كيفية الحفاظ على تجربة تطور الإشريكية القولونية طويلة الأجل (LTEE) من خلال إجراء عمليات النقل اليومية والتجميد الدوري وكيفية إجراء فحوصات المنافسة لقياس تحسينات اللياقة البدنية في البكتيريا المتطورة. يمكن أن تكون هذه الإجراءات بمثابة نموذج للباحثين الذين يبدأون تجارب التطور الميكروبي الخاصة بهم.
Abstract
تابعت تجربة التطور طويلة المدى (LTEE) اثني عشر مجموعة من الإشريكية القولونية حيث تكيفت مع بيئة مختبرية بسيطة لأكثر من 35 عاما و 77000 جيل بكتيري. يجسد الإعداد والإجراءات المستخدمة في LTEE طرقا موثوقة وقابلة للتكرار لدراسة التطور الميكروبي. في هذا البروتوكول ، نصف أولا كيف يتم نقل مجموعات LTEE إلى وسط جديد واستزراعها كل يوم. بعد ذلك ، نصف كيف يتم فحص مجموعات LTEE بانتظام بحثا عن علامات التلوث المحتملة وأرشفتها لتوفير “سجل أحفوري” دائم مجمد لدراسته لاحقا. تم تصميم ضمانات متعددة مضمنة في هذه الإجراءات لمنع التلوث ، واكتشاف المشكلات المختلفة عند حدوثها ، والتعافي من الاضطرابات دون حدوث إعاقة ملحوظة لتقدم التجربة. إحدى الطرق التي يتم بها مراقبة الإيقاع العام وطبيعة التغيرات التطورية في LTEE هي قياس اللياقة التنافسية للسكان والسلالات من التجربة. نصف كيفية إجراء فحوصات المنافسة في الثقافة المشتركة ونوفر كلا من جدول البيانات وحزمة R (fitnessR) لحساب اللياقة النسبية من النتائج. على مدار LTEE ، تغيرت سلوكيات بعض السكان بطرق مثيرة للاهتمام ، ووفرت التقنيات الجديدة مثل تسلسل الجينوم الكامل سبلا إضافية للتحقيق في كيفية تطور السكان. نختتم بمناقشة كيفية تحديث إجراءات LTEE الأصلية لاستيعاب هذه التغييرات أو الاستفادة منها. سيكون هذا البروتوكول مفيدا للباحثين الذين يستخدمون LTEE كنظام نموذجي لدراسة الروابط بين التطور وعلم الوراثة والبيولوجيا الجزيئية وبيولوجيا الأنظمة والبيئة. على نطاق أوسع ، يوفر LTEE نموذجا مجربا وصحيحا لأولئك الذين يبدأون تجارب التطور الخاصة بهم مع الميكروبات والبيئات والأسئلة الجديدة.
Introduction
في فبراير من عام 1988 ، قام ريتشارد لينسكي بتلقيح اثني عشر قارورة تحتوي على وسط نمو محدود الجلوكوز مع مزارع نسيلية من الإشريكية القولونية في جامعة كاليفورنيا ، إيرفين1. في اليوم التالي ، نقل 1٪ من الثقافة من كل قارورة إلى مجموعة من القوارير الجديدة التي تحتوي على وسط نمو جديد. سمح هذا التخفيف بنسبة 1: 100 لمجموعات البكتيريا بالتوسع 100 ضعف قبل استنفاد الجلوكوز المتاح ، وهو ما يعادل حوالي 62/3 أجيال من انقسامات الخلايا. تم تكرار هذا الإجراء في اليوم التالي وكان كل يوم منذ ذلك الحين ، مع بعض الانقطاعات. استمرت عمليات النقل اليومية هذه ، حتى مع نقل التجربة ، أولا إلى جامعة ولاية ميشيغان في عام 1992 ، ثم إلى جامعة تكساس في أوستن في عام 2022. طوال الوقت ، ولدت الطفرات الجديدة باستمرار تباينا وراثيا في مجموعات الإشريكية القولونية هذه ، وأدى الانتقاء الطبيعي إلى خلايا متطورة تتفوق على أسلافها.
صمم لينسكي هذه التجربة ، المعروفة الآن باسم تجربة التطور طويلة المدى (LTEE) ، للتحقيق في ديناميكيات التطور وتكراره. للإجابة على هذه الأسئلة ، قام بتضمين العديد من الميزات المهمة في تصميم الإعداد التجريبي وبروتوكولاته2. واحدة من هذه الميزات كانت الاختيار الدقيق لكائن نموذجي. بدأت جميع المجموعات السكانية الاثني عشر الأصلية من مستعمرات فردية تشترك في سلف مشترك مباشر ، سلالة الإشريكية القولونية B REL606. تم اختيار هذه السلالة لأنها كانت شائعة الاستخدام بالفعل في إعدادات المختبر ، وتم استنساخها بشكل لا جنسي تماما ، ولا تحتوي على بلازميدات أو نبوءات سليمة 3,4 – وكلها تجعل دراسة تطورها أكثر بساطة. كان الخيار الآخر الذي بسط التجربة هو استخدام تركيز منخفض جدا من الجلوكوز في وسط النمو للحد من كثافة الخلايا في كل دورق بعد النمو. كان الهدف من استخدام كثافة الخلايا المنخفضة هو تسهيل تحليل التغيرات في اللياقة السكانية عن طريق تقليل احتمالية تطور التفاعلات البيئية داخل السكان (على سبيل المثال ، عن طريق التغذية المتبادلة)5.
REL606 غير قادر على استخدام أرابينوز كمصدر للكربون والطاقة (Ara−) بسبب طفرة نقطية في جين araA. قبل بدء LTEE ، تم عزل متحولة عفوية مع تسلسل araA مستعاد ، المسمى REL607 ، من REL6066. REL607 قادر على النمو على أرابينوز (آرا +). تم استخدام REL606 لبدء ستة من مجموعات LTEE ، وتم استخدام REL607 لبدء الستة الآخرين. أرابينوز غير موجود في وسط النمو المستخدم خلال LTEE ، لذلك يتصرف REL607 مثل REL606 في ظل هذه الظروف. ومع ذلك ، عند الطلاء على أجار رباعي أرابينوز (TA) ، تشكل خلايا Ara− و Ara + مستعمرات حمراء وبيضاء ، على التوالي. هذه الطريقة للتمييز بين سلالتين من الإشريكية القولونية وأحفادهما مفيدة للغاية. يمكن استخدامه للكشف عن التلوث المتبادل بين مجموعات LTEE. كما أنه يساعد في قياس لياقة سلالة Ara− أو السكان بالنسبة إلى سلالة Ara+ عندما يتنافسون ضد بعضهم البعض. يتم قياس اللياقة من خلال إنشاء ثقافة مشتركة للمنافسين المميزين بشكل معاكس ثم مراقبة كيفية تغير ترددات المستعمرات الحمراء والبيضاء (التي يتم الحصول عليها عن طريق نشر تخفيفات الثقافة على لوحات TA) بين وقت خلط المنافسين في البداية وبعد دورة نمو واحدة أو أكثر في ظل نفس الظروف مثل LTEE. سيزداد تمثيل نوع الخلية الأكثر ملاءمة خلال كل دورة نمو.
ميزة أخرى حاسمة ل LTEE هي أن عينات من السكان المتطورين يتم أرشفتها بشكل دوري. عند مزجها مع مادة واقية من البرودة مثل الجلسرين ، يمكن تجميد خلايا الإشريكية القولونية وإحيائها لاحقا7. كجزء من بروتوكول LTEE ، كل يوم 75 (أي ما يعادل ما يقرب من 500 جيل) ، يتم خلط جزء من كل مجموعة لم يتم نقلها إلى قارورة جديدة مع الجلسرين ، وتقسيمها بين قوارير متعددة ، وتخزينها في الثلاجة. مكن هذا “السجل الأحفوري” المجمد الباحثين من إجراء الدراسات الأولى ل LTEE ، حيث قاموا بإحياء مجموعات E. coli المتطورة من نقاط زمنية مختلفة وتنافسوا عليها مقابل سلالات الأجداد لتتبع مدى سرعة زيادة اللياقةالبدنية 1. تم إعادة قياس تطور اللياقة البدنية بشكل دوري حيث تم الحفاظ على المزيد من “طبقات” “السجل الأحفوري” المجمد. الاستنتاج العام من هذه القياسات هو أن اللياقة البدنية تستمر في التحسن في LTEE حتى يومنا هذا ، حتى بعد أجيال عديدة من التطور في نفس البيئة8،9،10.
ما الذي سمح ل LTEE بالاستمرار لفترة طويلة؟ العديد من الميزات نفسها التي مكنت من طرح أسئلتها الأصلية والإجابة عليها كانت أيضا بمثابة تدابير أمان وأمان ضد الاضطرابات الحتمية بسبب سوء الحظ والخطأ البشري والأحداث العالمية. كل يوم ، عندما يتم نقل الثقافات إلى وسط نمو جديد ، يتناوب الباحث الذي يقوم بعمليات النقل بين مجموعات Ara− و Ara+ . بعد ذلك ، عندما يتم تجميد السكان ، يمكن طلاءهم على أجار انتقائي ومؤشر للتحقق مما إذا كانت أي مجموعات “مجاورة” قد تلوثت أو اختلطت عن طريق الخطأ (على سبيل المثال ، المستعمرات البيضاء موجودة في مجموعة يجب أن تشكل مستعمرات حمراء فقط) أو ملوثة بميكروبات غريبة (على سبيل المثال ، مورفولوجيا مستعمرة غير متوقعة أو كثافة خلية). في حالة تعرض السكان للخطر ، يمكن إحياء سلفه من الثلاجة ونقله في مكانه. وبالتالي ، فإن علامات Ara والأرشيف المجمد تخدم أغراضا مزدوجة كموارد تجريبية وتدابير سلامة.
نظرا لأن تاريخها محفوظ جيدا ويمكن الوصول إليه بسهولة ، فقد تمت دراسة عينات LTEE باستخدام تقنيات لم تكن موجودة عندما بدأت التجربة. على سبيل المثال ، تم استخدام تسلسل الجينوم الكامل لفحص ديناميكيات الطفرات في مجموعات LTEE11،12،13،14،15 ، وتم استخدام علم النسخ والتنميط الريبوسومي لفحص التغيرات في التعبير الجيني16،17. تم استخدام الأدوات الجينية لإعادة بناء السلالات التي تختلف عن طريق الطفرات المفردة أو مجموعات من العديد من الطفرات المتطورة لفهم آثارها على اللياقة البدنية والأنماط الظاهرية المختلفة18،19،20،21. يتم تجديد العينات من “السجل الأحفوري” المجمد بسهولة بحيث يمكن شحن أجزاء من أو نسخ كاملة من تاريخ التجربة إلى مختبرات أخرى. توجد عينات LTEE الآن في جميع القارات باستثناء القارة القطبية الجنوبية ، ويتم دراستها من قبل باحثين أصغر سنا من التجربة نفسها. كما كانت الطرق القوية ل LTEE وعينات وسلالات الإشريكية القولونية المتطورة من سجلها التاريخي بمثابة نقاط انطلاق لتجارب التطور التي تدرس الأسئلة والبيئات الأخرى 22،23،24،25،26،27،28،29.
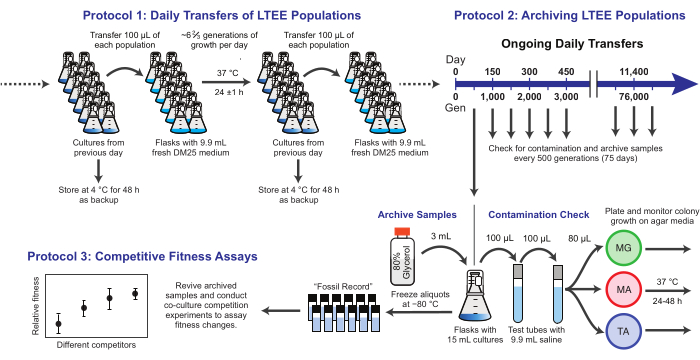
الشكل 1: نظرة عامة على إجراءات LTEE. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
هنا ، نوضح ثلاثة بروتوكولات أساسية مستخدمة في تجربة تطور الإشريكية القولونية طويلة المدى (الشكل 1). وصفنا: (1) كيفية إجراء عمليات النقل اليومية ، (2) كيفية أرشفة عينات السكان والعزلات النسيلية ، و (3) كيفية إجراء وتحليل مقايسات المنافسة في الثقافة المشتركة لقياس اختلافات اللياقة البدنية. نأمل أن تعزز هذه البروتوكولات الاستخدام المستمر لموارد LTEE وإبلاغ تصميم تجارب التطور الميكروبي الجديدة.
Protocol
Representative Results
Discussion
المرونة طويلة الأجل ل LTEE وأساليبها
تجربة التطور طويل الأجل للإشريكية القولونية (LTEE) هي الآن في عقدها الرابع. بالنسبة لتجربة التطور الميكروبي لأي مدة ، من الأهمية بمكان الحفاظ على بيئة قابلة للتكرار ، وتجنب التلوث ، وأرشفة العينات ، وقياس اللياقة بدقة. يوضح LTEE العديد من الاستراتيجيات التي تم اختبارها عبر الزمن لتحقيق هذه الأهداف ، بما في ذلك استخدام قوارير مهتزة جيدا تخلق بيئة متجانسة ووسط نمو محدد كيميائيا يدعم كثافة الخلايا المنخفضة. علاوة على ذلك ، يستخدم LTEE سلالات أسلاف تختلف في علامة جينية تعطي نمطا ظاهريا (لون مستعمرة) يسهل فحصه ومحايدا بشكل انتقائي في بيئة التطور. توفر ميزة التصميم التجريبي هذه وسيلة لتحديد التلوث الداخلي والخارجي وتسهل قياس اللياقة. ومع ذلك ، لم تثبت جميع الإجراءات والضمانات التي استخدمها LTEE منذ عام 1988 أنها قوية بنفس القدر. أصبحت بعض الطرق التي كانت موثوقة عندما بدأت LTEE أقل فعالية مع تطور مجموعات الإشريكية القولونية . لحسن الحظ ، يمكن الآن زيادة هذه الأساليب الإشكالية أو استبدالها باستخدام التقنيات التي تم تطويرها منذ بداية التجربة.
الكشف عن التلوث
يعد اكتشاف التلوث أمرا بالغ الأهمية ل LTEE. يمكن أن يكون التلوث من نوعين: بين مجموعات LTEE (التلوث المتبادل) ومع الميكروبات من البيئة (التلوث الخارجي). بالنسبة للجزء الأكبر ، فإن الاستخدام الدقيق لتقنيات التعقيم والاهتمام الوثيق أثناء إعداد الوسائط وعمليات النقل اليومية يمنع كلا النوعين من التلوث ، لكنهما يحدثان. في وقت مبكر من التجربة ، يمكن استخدام الطلاء على TA agar للكشف عن حالات التلوث المتبادل لأن عمليات النقل كانت دائما بالتناوب بين مجموعات Ara− و Ara+. كان المقصود أيضا أن تكون بصمة حساسية ومقاومة هذه الإشريكية القولونية لبعض البكتيريا ميزة تصميم يمكن أن تميز مجموعات LTEE عن سلالات مختبر الإشريكية القولونية شائعة الاستخدام التي قد تلوثها4. ومع ذلك ، أصبحت هذه العلامات الجينية غير موثوقة مع تقدم التجربة (على سبيل المثال ، لم تعد بعض المجموعات السكانية تشكل مستعمرات على TA agar)10,35. لحسن الحظ ، تباعدت المجموعات وراثيا لأنها شهدت تاريخا تطوريا منفصلا خلال التجربة ، مما أدى إلى إنشاء علامات جينية جديدة يمكن استخدامها الآن للكشف عن التلوث المتبادل. على سبيل المثال ، طور كل مجتمع مجموعة مجموعة فريدة من الطفرات في جينات pykF و nadR 14،36،37. نقوم أحيانا بتضخيم تفاعل البوليميراز المتسلسل وتسلسل سانجر هذين الجينين لاختبار ما إذا كانت المستعمرات ذات الأشكال أو الألوان غير العادية ناتجة عن التلوث المتبادل. مع استمرار انخفاض تكاليف الجينوم الكامل وتسلسل السكان بالكامل ، قد يكون التسلسل الروتيني لمجموعات LTEE ممكنا قريبا ، مما يوفر فرصا جديدة لمراقبتها بحثا عن علامات التلوث.
قياس اللياقة التنافسية
حالة أخرى تجاوزت فيها LTEE أساليبها الأصلية هي أن لياقة الإشريكية القولونية المتطورة قد زادت في البيئة التجريبية لدرجة أنه لم يعد بإمكان المرء قياس لياقة سكان اليوم بشكل مباشر بالنسبة لأسلافهم باستخدام البروتوكول الموصوف هنا. تتفوق المجموعات السكانية المتطورة على الأسلاف لدرجة أنه لم يتبق سوى عدد قليل من مستعمرات الأسلاف أو عدم عدمها بعد منافسة استمرت يوما واحدا. تتمثل إحدى طرق التعامل مع هذا الاختلاف الكبير في اللياقة في استخدام نسب بدء غير متساوية للسلالات ، وترجيح الأحجام الأولية المختلطة نحو المنافس الأقل ملاءمة (على سبيل المثال ، سلف 90 ميكرولتر ومنافس متطور 10 ميكرولتر). تتمثل الطريقة الثانية في تحديد استنساخ Ara− المتطور الذي يتمتع بلياقة أعلى من سلف LTEE ، وعزل متحولة Ara + العفوية منه عن طريق الاختيار على MA agar ، ثم التحقق من أن سلالة العاكس لها نفس اللياقة مثل أصلها باستخدام اختبار المنافسة 6,38. يمكن بعد ذلك استخدام زوج Ara− / Ara + الجديد هذا كمجموعة من سلالات المنافسين الشائعة بدلا من REL606 / REL607. من الناحية المثالية ، سيكون لاستنساخ Ara− المتطور الذي تم اختياره كمنافس مشترك (وعائده Ara+ ) لياقة متوسطة بالنسبة لجميع السلالات ذات الاهتمام في التجربة. على مدار أول 50000 جيل من LTEE ، لم ينتج هذان النهجان (باستخدام نسب بدء غير متكافئة أو منافس مشترك) قياسات لياقة مختلفة بشكل هادف مقابل النهج النموذجي39.
هذه التعديلات على بروتوكول المنافسة تجعل بعض الافتراضات المبسطة التي قد لا تكون صحيحة دائما. الأول هو أن قياسات اللياقة البدنية متعدية. وهذا يعني أننا إذا تنافسنا بين مجموعتين سكانيتين مقابل سلالة منافسة مشتركة بشكل منفصل ، فيمكننا استنتاج اللياقة النسبية للمجموعتين لبعضهما البعض. تم العثور على هذه العلاقة لتكون صحيحة بالنسبة ل LTEE40 ، بالنسبة للجزء الأكبر ، ولكنها ليست للتجارب الأخرى41. يمكن أن يكون أحد أسباب هذا التناقض هو تطور تأثيرات اللياقة البدنية السلبية المعتمدة على التردد. يحدث هذا الموقف عندما تتنافس سلالات معزولة من سلالتين مختلفتين متباعدتين من السكان A − 2 من LTEE ضد بعضها البعض19,42. لكل منها ميزة عندما تكون نادرة ، بسبب التغذية المتقاطعة ، مما يستقر في تعايشها. تشير بيانات التسلسل التي تظهر تعايشا طويل الأمد للسلالات مع مجموعات مختلفة من الطفرات إلى أن تفاعلات مماثلة ربما نشأت أيضا في مجموعات LTEE الأخرى14,43 ، على الرغم من أنه ليس من الواضح ما إذا كانت قوية بما يكفي لتغيير تقديرات اللياقة بشكل ملحوظ. أخيرا ، فإن تطور النمو الهوائي على السيترات في السكان A − 3 من LTEE32 يعني أن لياقة هذه الخلايا تتضمن الآن استخدام مورد “خاص” عندما يتم التنافس مع الخلايا التي لا يمكنها استخدام السيترات ، مما يعقد تفسير هذه النتائج. على الرغم من هذه الاستثناءات ، فإن استخدام تركيز منخفض للجلوكوز وبيئة مهتزة جيدا قد بسط بلا شك إجراء مقارنات للياقة البدنية بين سلالات LTEE والسكان.
في الأجيال اللاحقة ، لم تعد بعض مجموعات LTEE تشكل مستعمرات على TA agar ، مما يجعل إجراء تجارب المنافسة باستخدام حتى البروتوكولات المعدلة أمرا صعبا أو مستحيلا10. يمكن استخدام طرق بديلة لا تتطلب نمو مستعمرة لتحديد التمثيل النسبي لمنافسين ، مثل FREQ-seq الذي يستخدم تسلسل الجيل التالي لحساب نسبة القراءات التي تحتوي على أليلين بديلين في amplicon44. يمكن استخدام هذه الطريقة أو طريقة مماثلة مع أليلات آرا أو مع الطفرات المتطورة حديثا ، مثل تلك الموجودة في pykF و nadR ، مقابل تسلسل الأسلاف. يمكن أيضا استخدام إجراء التعديلات الجينية التي تقدم أنواعا أخرى من العلامات المحايدة لقياس اللياقة النسبية. على سبيل المثال ، تم إدخال جينات البروتين الفلوري في كروموسومات الخلايا في تجارب فرع LTEE بحيث يمكن حساب المنافسين باستخدام قياس التدفقالخلوي 45. هناك نهج آخر ، يفتح إمكانية خلط أكثر من سلالتين معا في نفس قارورة المنافسة ، وهو إدخال الرموز الشريطية التي يمكن تضخيمها وتسلسلها في جينومات المنافسين المختلفين. تم استخدام هذا النهج لتتبع النسب في تجارب التطور46. يمكن لكل من قياس التدفق الخلوي وتسلسل الباركود أن يقيس بدقة النسب الأكثر تطرفا لسلالتين مقابل عد المستعمرات (لأنه يمكنهم الاستعلام عن > 10000 خلية / جينوم مقابل < 500 التي يمكن عدها على لوحة أجار) ، لذا فإن استخدام هذه الطرق يعد أيضا بزيادة النطاق الديناميكي من حيث اختلافات اللياقة التي يمكن قياسها بالنسبة إلى منافس مشترك.
تصاميم بديلة لتجارب التطور الميكروبي طويلة الأجل
على الرغم من كل فضائلها ، فإن LTEE ليست مثالية. بعض جوانب تصميمه تجعله كثيف العمالة وعرضة للخطأ البشري. على سبيل المثال ، يجب أن يأتي الباحث كل يوم إلى المختبر والماصة بين قوارير Erlenmeyer لمواصلة التجربة. يمكن أن تشكل تجارب المنافسة أيضا عقبات لوجستية شاقة ، بالنظر إلى أن متطلبات الأواني الزجاجية المعقمة ، والوسائط ، ومساحة الحاضنة ، وعد المستعمرات تتصاعد بسرعة عندما يتم اختبار عدد صغير من المنافسين بتكرار متواضع. غالبا ما يسألنا لماذا لا نستفيد من أنظمة أتمتة المختبرات ، مثل روبوتات السحب التي تعمل على 96 صفيحة دقيقة ، أو أنظمة الاستزراع المستمر ، مثل chemostats أو turbidostats. الجواب بسيط: LTEE ، إلى حد ما ، سجين لتاريخه الطويل. لا نجرؤ على الانحراف عن ثقافات 10 مل تهتز بسرعة محددة في قوارير Erlenmeyer سعة 50 مل لأن هذا من شأنه أن يخاطر بتغيير التجربة بشكل جذري. سيتم تغيير الجوانب الدقيقة للبيئة التي كان هؤلاء السكان يتكيفون معها لعقود (على سبيل المثال ، كمية التهوية) في الصفائح الدقيقة أو أنظمة الاستزراع المستمر. قد يكون عنق الزجاجة السكاني في كل عملية نقل مختلفا أيضا (أصغر في الصفائح الدقيقة ، على سبيل المثال) ، مما يغير الديناميات التطورية. باختصار ، فإن الانحراف عن الأساليب الموضحة هنا من شأنه أن يجعل LTEE تجربة مختلفة ، أو على الأقل يخاطر بإدخال انقطاع من شأنه أن يعطل المسارات التطورية.
يجب على الباحثين الذين يصممون تجارب تطور جديدة النظر في هذه الطرق الأخرى لنشر المجموعات الميكروبية ، مع إدراك فوائدها وعيوبها المحتملة. يعد استخدام روبوتات السحب لنقل السكان في ألواح microwell أبسط من الناحية اللوجستية من بعض النواحي ويمكن أن يثبت أنه قوي جدا بسبب الأعداد الكبيرة من المجموعات المكررة التي يمكن نشرها بهذه الطريقة47،48،49. ومع ذلك ، فإن عمليات النقل الآلية في معظم الإعدادات الحالية لا تتم في ظل ظروف معقمة تماما ، مما يزيد من احتمال التلوث الخارجي. لمنع التلوث ، غالبا ما يتم استكمال وسط النمو بالمضادات الحيوية ، والتي تصبح سمة من سمات البيئة التي تؤثر على التطور. كما أن عمليات النقل في ألواح microwell أكثر عرضة لأحداث التلوث المتبادل. أخيرا ، تميل بيئة ألواح microwell – خاصة إذا لم تهتز – إلى اختيار نمو الجدار والتجميع والظواهر الأخرى التي يمكن أن تعقد التطور من خلال إنشاء منافذ متعددة في بئر واحدة. من المرجح أن يؤدي استخدام الوسائط الغنية أو التركيزات العالية من العناصر الغذائية للحفاظ على أحجام السكان الكبيرة في الآبار الصغيرة إلى تفاقم هذه التعقيدات. إذا ظهرت مثل هذه التفاعلات ، فإنها يمكن أن تجعل قياس وتفسير اللياقة البدنية أكثر صعوبة.
تشمل أنظمة الاستزراع المستمر للتطور الميكروبي chemostats ، حيث يتم ضخ وسط جديد باستمرار ويتم ضخ الثقافة ، و turbidostats ، حيث يتم تخفيف الثقافات بشكل دوري من خلال الاستشعار الآلي والضخ للحفاظ على الخلايا في حالة نمو مستمر. هذه الأنظمة مفيدة للغاية عندما يرغب المرء في نمذجة علم وظائف الأعضاء الميكروبية والتطور لأنها تتجنب انتقال الميكروبات بين النمو والمجاعة عن طريق الاحتفاظ بها في بيئة تحتوي دائما على العناصر الغذائية50. يمكن للمرء حتى إضافة أجهزة استشعار تقوم بإجراء قياسات في الوقت الفعلي للكثافة البصرية ، واستهلاك O2 ، ودرجة الحموضة ، والجوانب الأخرى لبيئة الثقافة ونموها. ومع ذلك ، تتطلب أنظمة الثقافة المستمرة الحالية إما شراء معدات باهظة الثمن أو خبرة متخصصة لبناء إعدادات مخصصة51،52،53،54. كما أن نمو الجدار، الذي تفلت فيه الخلايا من التخفيف عن طريق الالتصاق بغرفة الثقافة، يفسد الديناميات التطورية في أنظمة الاستزراع المستمر ما لم يتم تعقيمها بشكل دوري. بسبب هذه القيود ، كانت معظم تجارب تطور chemostat و turbidostat حتى الآن محدودة المدة و / أو تضمنت عددا قليلا نسبيا من المجموعات المتطورة بشكل مستقل مقارنة بتجارب تطور النقل المتسلسل.
استنتاج
تعتبر الطرق التي نوضحها هنا ل LTEE ضرورية لدراسة سجلها التاريخي الفريد ومواصلة التطور المفتوح لمجموعات الإشريكية القولونية هذه. كما أنها توفر نقطة انطلاق للآخرين الذين يفكرون في تجارب تطور جديدة قد تستفيد من أتمتة المختبرات أو تضيف عناصر مختلفة من التعقيد الموجود في البيئات الطبيعية التي تم حذفها عمدا من LTEE. منذ عام 1988 ، ازدهر التطور التجريبي كمجال. خلال هذا الوقت ، أظهر الباحثون في المختبرات في جميع أنحاء العالم المرونة الهائلة لهذا النهج لدراسة التطور والابتكار من خلال تقديم تصميمات تجريبية إبداعية ومراقبة النتائج باستخدام تقنيات جديدة. لا تمثل أساليب LTEE نقطة نهاية ، لكننا نأمل أن تستمر في إلهام وتوفير أساس للمجال في المستقبل البعيد.
Divulgazioni
The authors have nothing to disclose.
Acknowledgements
نشكر ريتشارد لينسكي والعديد من الباحثين الذين درسوا وساهموا في الحفاظ على تجربة التطور طويلة المدى مع الإشريكية القولونية ، بما في ذلك على وجه الخصوص نيرجا هاجيلا. يتم دعم LTEE حاليا من قبل المؤسسة الوطنية للعلوم (DEB-1951307).
Materials
| 2,3,5-Triphenyltetrazolium chloride (TTC) | Sigma-Aldrich | T8877 | |
| 20 mL Glass Beaker | Sigma-Aldrich | CLS100020 | |
| 50 mL Erlenmeyer Flasks | Sigma-Aldrich | CLS498050 | |
| Agar | Sigma-Aldrich | A1296 | |
| Ammonium Sulfate | Sigma-Aldrich | AX1385 | |
| Antifoam | Sigma-Aldrich | A5757 | |
| Arabinose | Sigma-Aldrich | A3256 | |
| Freezer Box (2") | VWR | 82007-142 | |
| Freezer Box (3") | VWR | 82007-144 | |
| Freezer Box Cell Divider (49-place) | VWR | 82007-150 | |
| Freezer Box Cell Divider (81-place) | VWR | 82007-154 | |
| Freezer Vials (1/2-Dram) | VWR | 66009-816 | |
| Freezer Vials (2-Dram) | VWR | 66010-560 | |
| Glucose | Sigma-Aldrich | G8270 | |
| Glycerol | Fisher Scientific | G33 | |
| Magnesium Sulfate | Sigma-Aldrich | M7506 | |
| Metal Tray | Winco | SPJP-202 | |
| Petri Dish | Fisher Scientific | FB0875712 | |
| Potassium Phosphate Dibasic Trihydrate | Sigma-Aldrich | P5504 | |
| Potassium Phosphate Monobasic | Sigma-Aldrich | P5379 | |
| Sodium Chloride | Sigma-Aldrich | M7506 | |
| Sodium Citrate Tribasic Dihydrate | Sigma-Aldrich | C7254 | |
| Test Tube Cap (18mm) | VWR | 10200-142 | |
| Test Tube Rack (18mm, steel) | Adamas-Beta | N/A | Test Tube Racks Stainless Steel Grid Arrangement 72 Holes (17-19 mm) |
| Test Tubes (18 x 150 mm) | VWR | 47729-583 | |
| Thiamine, Hydrochloride | Millipore | 5871 | |
| Tryptone | Gibco | 211705 | |
| Yeast Extract | Gibco | 212750 |
Riferimenti
- Lenski, R. E., Rose, M. R., Simpson, S. C., Tadler, S. C. Long-term experimental evolution in Escherichia coli. I. Adaptation and divergence during 2,000 generations. The American Naturalist. 138 (6), 1315-1341 (1991).
- Fox, J. W., Lenski, R. E. From here to eternity-the theory and practice of a really long experiment. PLoS Biology. 13 (6), e1002185 (2015).
- Daegelen, P., Studier, F. W., Lenski, R. E., Cure, S., Kim, J. F. Tracing ancestors and relatives of Escherichia coli B, and the derivation of B strains REL606 and BL21(DE3). Journal of Molecular Biology. 394 (4), 634-643 (2009).
- Studier, F. W., Daegelen, P., Lenski, R. E., Maslov, S., Kim, J. F. Understanding the differences between genome sequences of Escherichia coli B strains REL606 and BL21(DE3) and comparison of the E. coli B and K-12 genomes. Journal of Molecular Biology. 394 (4), 653-680 (2009).
- Barrick, J. E., Lenski, R. E. Genome dynamics during experimental evolution. Nature Reviews Genetics. 14 (12), 827-839 (2013).
- Lenski, R. E. Experimental studies of pleiotropy and epistasis in Escherichia coli. II. Compensation for maladaptive pleiotropic effects associated with resistance to virus T4. Evolution. 42 (3), 425-432 (1988).
- Calcott, P. H., Gargett, A. M. Mutagenicity of freezing and thawing. FEMS Microbiology Letters. 10 (2), 151-155 (1981).
- Lenski, R. E., Travisano, M. Dynamics of adaptation and diversification: a 10,000-generation experiment with bacterial populations. Proceedings of the National Academy of Sciences of the United States of America. 91 (15), 6808-6814 (1994).
- Wiser, M. J., Ribeck, N., Lenski, R. E. Long-term dynamics of adaptation in asexual populations. Science. 342 (6164), 1364-1367 (2013).
- Lenski, R. E., et al. Sustained fitness gains and variability in fitness trajectories in the long-term evolution experiment with Escherichia coli. Proceedings of the Royal Society B: Biological Sciences. 282 (1821), 20152292 (2015).
- Barrick, J. E., et al. Genome evolution and adaptation in a long-term experiment with Escherichia coli. Nature. 461 (7268), 1243-1247 (2009).
- Blount, Z. D., Barrick, J. E., Davidson, C. J., Lenski, R. E. Genomic analysis of a key innovation in an experimental Escherichia coli population. Nature. 489 (7417), 513-518 (2012).
- Tenaillon, O., et al. Tempo and mode of genome evolution in a 50,000-generation experiment. Nature. 536 (7615), 165-170 (2016).
- Good, B. H., McDonald, M. J., Barrick, J. E., Lenski, R. E., Desai, M. M. The dynamics of molecular evolution over 60,000 generations. Nature. 551 (7678), 45-50 (2017).
- Consuegra, J., et al. Insertion-sequence-mediated mutations both promote and constrain evolvability during a long-term experiment with bacteria. Nature Communications. 12 (1), 980-980 (2021).
- Cooper, T. F., Rozen, D. E., Lenski, R. E. Parallel changes in gene expression after 20,000 generations of evolution in Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America. 100 (3), 1072-1077 (2003).
- Favate, J. S., Liang, S., Cope, A. L., Yadavalli, S. S., Shah, P. The landscape of transcriptional and translational changes over 22 years of bacterial adaptation. eLife. 11, e81979 (2022).
- Khan, A. I., Dinh, D. M., Schneider, D., Lenski, R. E., Cooper, T. F. Negative epistasis between beneficial mutations in an evolving bacterial population. Science. 332 (6034), 1193-1196 (2011).
- Plucain, J., et al. Epistasis and allele specificity in the emergence of a stable polymorphism in Escherichia coli. Science. 343 (6177), 1366-1369 (2014).
- Quandt, E. M., Deatherage, D. E., Ellington, A. D., Georgiou, G., Barrick, J. E. Recursive genomewide recombination and sequencing reveals a key refinement step in the evolution of a metabolic innovation in Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America. 111 (6), 2217-2222 (2014).
- Leon, D., D’Alton, S., Quandt, E. M., Barrick, J. E. Innovation in an E. coli evolution experiment is contingent on maintaining adaptive potential until competition subsides. PLoS Genetics. 14 (4), e1007348 (2018).
- Bennett, A. F., Lenski, R. E., Mittler, J. E. Evolutionary adaptation to temperature. I. Fitness responses of Escherichia coli to changes in its thermal environment. Evolution. 46 (1), 16-30 (1992).
- Kibota, T. T., Lynch, M. Estimate of the genomic mutation rate deleterious to overall fitness in E. coli. Nature. 381 (6584), 694-696 (1996).
- Friesen, M. L., Saxer, G., Travisano, M., Doebeli, M. Experimental evidence for sympatric ecological diversification due to frequency-dependent competition in Escherichia coli. Evolution. 58 (2), 245-260 (2004).
- Cooper, T. F. Recombination speeds adaptation by reducing competition between beneficial mutations in populations of Escherichia coli. PLoS Biology. 5 (9), e225 (2007).
- Cooper, T. F., Lenski, R. E. Experimental evolution with E. coli in diverse resource environments. I. Fluctuating environments promote divergence of replicate populations. BMC Evolutionary Biology. 10, 11 (2010).
- Quan, S., et al. Adaptive evolution of the lactose utilization network in experimentally evolved populations of Escherichia coli. PLoS Genetics. 8 (1), e1002444 (2012).
- Deatherage, D. E., Kepner, J. L., Bennett, A. F., Lenski, R. E., Barrick, J. E. Specificity of genome evolution in experimental populations of Escherichia coli evolved at different temperatures. Proceedings of the National Academy of Sciences of the United States of America. 114 (10), E1904-E1912 (2017).
- Izutsu, M., Lake, D. M., Matson, Z. W. D., Dodson, J. P., Lenski, R. E. Effects of periodic bottlenecks on the dynamics of adaptive evolution in microbial populations. BioRixv. , 4457 (2021).
- Chavarria-Palma, J. E., Blount, Z. D., Barrick, J. E. . LTEE Media Recipes. , (2022).
- Barrick, J. E., Lake, D. M. fitnessR: fitnessR-v1.0.0. barricklab. , (2023).
- Blount, Z. D., Borland, C. Z., Lenski, R. E. Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America. 105 (23), 7899-7906 (2008).
- Grant, N. A., Magid, A. A., Franklin, J., Dufour, Y., Lenski, R. E. Changes in cell size and shape during 50,000 generations of experimental evolution with Escherichia coli. Journal of Bacteriology. 203 (10), 22 (2021).
- Wickham, H. . ggplot2: Elegant Graphics for Data Analysis. , (2016).
- Meyer, J. R., et al. Parallel changes in host resistance to viral infection during 45,000 generations of relaxed selection. Evolution. 64 (10), 3024-3034 (2010).
- Woods, R., Schneider, D., Winkworth, C. L., Riley, M. A., Lenski, R. E. Tests of parallel molecular evolution in a long-term experiment with Escherichia coli. Proceedings of the National Academy of Sciences of the United States of America. 103 (24), 9107-9712 (2006).
- . LTEE-Ecoli: genomics resources for the Long-Term Evolution Experiment with Escherichia coli Available from: https://github.com/barricklab/LTEE-Ecoli (2022)
- Izutsu, M., Lenski, R. E. Experimental test of the contributions of initial variation and new mutations to adaptive evolution in a novel environment. Frontiers in Ecology and Evolution. 10, 958406 (2022).
- Wiser, M. J., Lenski, R. E. A comparison of methods to measure fitness in Escherichia coli. PLoS One. 10 (5), 0126210 (2015).
- de Visser, J. A. G. M., Lenski, R. E. Long-term experimental evolution in Escherichia coli. XI. Rejection of non-transitive interactions as cause of declining rate of adaptation. BMC Evolutionary Biology. 2 (1), 19 (2002).
- Paquin, C. E., Adams, J. Relative fitness can decrease in evolving asexual populations of S. cerevisiae. Nature. 306 (5941), 368-371 (1983).
- Rozen, D. E., Lenski, R. E. Long-Term Experimental Evolution in Escherichia coli. VIII. Dynamics of a balanced polymorphism. The American Naturalist. 155 (1), 24-35 (2000).
- Quandt, E. M., Gollihar, J., Blount, Z. D., Ellington, A. D., Georgiou, G., Barrick, J. E. Fine-tuning citrate synthase flux potentiates and refines metabolic innovation in the Lenski evolution experiment. eLife. 4, e09696 (2015).
- Chubiz, L. M., Lee, M. -. C., Delaney, N. F., Marx, C. J. FREQ-Seq: a rapid, cost-effective, sequencing-based method to determine allele frequencies directly from mixed populations. PLoS One. 7 (10), e47959 (2012).
- Gallet, R., Cooper, T. F., Elena, S. F., Lenormand, T. Measuring selection coefficients below 10-3: method, questions, and prospects. Genetica. 190 (1), 175-186 (2012).
- Levy, S. F., et al. Quantitative evolutionary dynamics using high-resolution lineage tracking. Nature. 519 (7542), 181-186 (2015).
- Lang, G. I., Botstein, D., Desai, M. M. Genetic variation and the fate of beneficial mutations in asexual populations. Genetica. 188 (3), 647-661 (2011).
- Frenkel, E. M., et al. Crowded growth leads to the spontaneous evolution of semistable coexistence in laboratory yeast populations. Proceedings of the National Academy of Sciences. 112 (36), 11306-11311 (2015).
- Jordt, H., et al. Coevolution of host-plasmid pairs facilitates the emergence of novel multidrug resistance. Nature Ecology and Evolution. 4 (6), 863-869 (2020).
- Gresham, D., Dunham, M. J. The enduring utility of continuous culturing in experimental evolution. Genomics. 104 (6), 399-405 (2014).
- Miller, A. W., Befort, C., Kerr, E. O., Dunham, M. J. Design and use of multiplexed chemostat arrays). Journal of Visualized Experiments. (72), e50262 (2013).
- Toprak, E., et al. Building a morbidostat: an automated continuous-culture device for studying bacterial drug resistance under dynamically sustained drug inhibition. Nature Protocols. 8 (3), 555-567 (2013).
- Wong, B. G., Mancuso, C. P., Kiriakov, S., Bashor, C. J., Khalil, A. S. Precise, automated control of conditions for high-throughput growth of yeast and bacteria with eVOLVER. Nature Biotechnology. 36 (7), 614-623 (2018).
- Ekkers, D. M., Branco Dos Santos, F., Mallon, C. A., Bruggeman, F., Van Doorn, G. S. The omnistat: A flexible continuous-culture system for prolonged experimental evolution. Methods in Ecology and Evolution. 11 (8), 932-942 (2020).
