Zebrafish Scale Regeneration In Toto and Ex Vivo Culture of Scales
Summary
This protocol describes the harvesting and visualization of elasmoid scales of zebrafish during in vivo regeneration. In addition, the ex vivo culture of these scales for up to 7 days after harvest is presented.
Abstract
Skeletal diseases are often complex in their etiology and affect millions of people worldwide. Due to the aging population, there is a need for new therapeutics that could ease the burden on healthcare systems. As these diseases are complex, it is difficult and expensive to accurately model bone pathophysiology in a lab setting. The challenge for the field is to establish a cost-effective, biologically relevant platform for modeling bone disease that can be used to test potential therapeutic compounds. Such a platform should ideally allow dynamic visualization of cell behaviors of bone-building osteoblasts and bone-degrading osteoclasts acting in their mineralized matrix environment. Zebrafish are increasingly used as models due to the availability of genetic tools, including transgenic reporter lines, and the fact that some skeletal tissues (including the scales) remain translucent to adulthood, allowing dynamic imaging options. Since zebrafish scales have both osteoblasts and osteoclasts and are highly abundant, they provide an easily accessible and abundantly available resource of independent bone units. Moreover, once removed, adult zebrafish scales fully regenerate, therefore offering a way to study the spatiotemporal growth of mineralized tissue in vivo. Here, we detail protocols for harvesting and tracking the regeneration of the scales. Lastly, a protocol for stable culture of scales ex vivo for a week and following the healing response after controlled damage to the mineralized matrix of the scale over time is also presented.
Introduction
Bone is a hard connective tissue that forms a major part of the skeleton, enabling locomotion and acting as a mineral reserve in the body. In order to maintain healthy bone, an exquisite balance between bone formation and degradation is essential via the coupled activity of osteoblasts (which are anabolic) and osteoclasts (which resorb bone). This balance is disrupted by aging or hormonal imbalance, often leading to bone fragility diseases such as osteoporosis1. Although existing drugs have been approved to target bone fragility diseases, many have side effects; therefore, there is a need for new therapeutics1. Thus, there remains a need for abundant sources of biologically relevant bone tissue that can be used to test potential therapeutic compounds.
Traditionally, rodent models and cell culture systems have been used to study bone biology. However, zebrafish are increasingly becoming another model of choice. While not a mammalian system, zebrafish offer certain advantages for bone research over rodents; these include their fecundity and the translucency of the larvae; even in adulthood, some skeletal tissues, including the scales and the fins, remain translucent, allowing high-resolution in vivo imaging and the increased availability of skeletal mutants2,3. Both the fins and scales of zebrafish are capable of complete regeneration after removal. Skeletal regeneration and injury repair of zebrafish fins have been studied extensively4,5, while zebrafish scales are a newer bone model in the field but offer advantages for ex vivo culture6.
Scales are highly abundant, with at least 300 scales on each fish that serve as a protective covering for the fish. Each scale is a small mineralized plate consisting of bone-forming osteoblasts and bone-resorbing osteoclasts of a collagen-rich skeletal matrix7. The ossification process of both zebrafish scales and human bones requires the differentiation of mesenchymal stem cells into osteoblasts to form the mineralized matrix. Zebrafish scales offer a great advantage for skeletal research with their strong regenerative ability that can be used to study bone regeneration and repair. However, despite the presence of both osteoblasts and osteoclasts, zebrafish scales lack osteocytes that are important for human bone remodeling and mechanosensation; the superficial location of the scales means that they can be easily removed with a pair of forceps. Upon removal of the scale, a cascade of events occurs, and scale regeneration begins8,9. There are various staining and imaging options available to visualize the activity of osteoblasts and osteoclasts and the mineralization of the scales, as shown in Figure 1. Additionally, the availability of many relevant fluorescent transgenic reporter lines of zebrafish means that one can visualize cell dynamics during regeneration7,10,11. This process allows one to gain more understanding of de novo bone formation by observing the early patterning of scale regeneration on the flank of the fish to study morphology, cellular activity, and genetic profiles of these regenerated scales. The biology of scale formation and regeneration has been well characterized. Importantly, scales can show a good predictive ability for therapeutically relevant compounds12 and treatment of fish with glucocorticoids leads to a scale that regenerates to show osteoporotic phenotypes13. The transcriptome of the regenerating scales shows that genes activated in scale regeneration are enriched for those linked to human skeletal diseases, further demonstrating their relevance as a model system6,14.
Finally, these scales can be cultured ex vivo for up to 7 days. Compared to cell line cultures that are typically composed of a single cell type, ex vivo scale culture provides in vitro bone study opportunities within its natural environment containing both osteoblasts and osteoclasts with its natural extracellular matrix8,12,15,16.
Scale culture also allows us to perform drug screening for novel osteoanabolic targets. The abundance of scales on the fish means that one can fill at least two plates of the 96-well plate from just a single fish, allowing for compound screening in a multiwell format where every single well contains one scale along with its natural niche of cells. Additionally, since the scales are thin, drug absorption is predictable12. In summary, the elasmoid scales of zebrafish have great potential in skeletal research and can help us gain more insight into the cellular events during bone formation and repair. Here, we describe protocols for harvesting scales to follow regeneration in vivo and culture the scales ex vivo.
Protocol
The University Animal Scientific Unit (ASU) is responsible for zebrafish care under the guidance of zebrafish husbandry guidelines. All procedures, including scale harvesting, live bone staining, and live imaging, were approved and performed under a UK Home Office Project License (PP4700996). For this manuscript, young adult transgenic zebrafish from the sp7:mCherry [Tg(osterix:mCherry-NTR)pd46] line was used17. The fish included both males and females of 4 months old.
1. In vivo scale regeneration
- Before starting the scale regeneration experiment, transfer zebrafish from their main tanks into individual tanks (see Table of Materials) and return to these individual tanks following imaging throughout the experiment, as shown in Figure 2. This is to ensure that all regenerating scales are those harvested for the experiment; group housing can lead to sporadic loss of scales due to injuries caused by other fish.
NOTE: The fish are individually placed to avoid fighting and subsequent scale loss between fish. - Record information for each experiment fish (i.e., genotype, age, and sex) according to the experiment design. Use a camera (smartphone cameras work fine) to take an image of each fish next to a ruler to record fish size/length and health condition.
- On the day of the experiment, anesthetize the zebrafish using 0.05% (v/v) tricaine methane sulfonate (MS222, see Table of Materials) by immersion, then place it on a Petri dish with a damp tissue to avoid movement of the fish during imaging or harvesting.
- Place the fish on its side (typically left flank is used here for consistency). Perform flank imaging using a stereomicroscope with appropriate magnification (typically, 2x, 4x, and/or 6.3x and 8x were used for the present study to capture both the overall regenerating area and the zoomed-in area for detailed observation).
NOTE: The flank imaging timepoints depend on the experimental purpose and choice of transgenic reporters. For example, to track osteoblasts during scale regeneration, the suggested timepoints are Day 0 (ontogenetic), 2 days post-harvest (dph), 4 dph, 7 dph, 10 dph, 14 dph, 21 dph. To reduce effects from repeated anesthesia, keep imaging timepoints to the fewest required for the experiment. - Harvest scales using fine tweezers and forceps under the stereomicroscope. Transfer the harvested scales into collection tubes for scale staining later.
NOTE: The maximum number of scales that can be harvested will depend on local ethical approvals that are in place. We typically harvest around 20 to 30 scales and collect them into individual collection tubes for scale staining later, such as ALP, von Kossa14. - Collect the scales into either a 1.5 mL or 2 mL collection tube. This depends on the staining technique used.
- For both ALP and von Kossa staining, transfer the scales into collection tubes containing deionized water, whereas for TRAP staining or immunohistochemistry, transfer the scales into collection tubes containing 4% PFA (fixing solution).
- Optional: Before transferring scales into tubes, capture images of individual harvested scales if required.
NOTE: Suggested scale harvest timepoints are day 0 (ontogenetic), day 10, and day 21 (regenerated scales).
2. Live bone staining
NOTE: Live skeletal staining can be performed either when experimental fish are not transgenic for fluorescent reporters or when carrying single-color transgenic reporters. Live staining can be used to complement transgenics; for example, one might use an osteoblast reporter such as sp7/osx in one color (e.g., GFP), then stain bone in red with Alizarin red (AR). AR and calcein green can be used for the same fish together; in this scenario, AR is used to label original or ontogenetic bone by binding to the mineralized matrix of aged bone, and calcein binds to calcium ions that are abundant in newly formed bone. For example, when staining an ontogenetic scale, AR can be visualized, but calcein green may be absent or minimal since ontogenetic scales have low levels of new bone formation.
- Perform live Alizarin Red (AR) staining.
- Prepare a bottle of 2x AR staining solution by mixing 0.5% (w/v) stock Alizarin solution (10 mL) with 10 mL of 1 M stock HEPES (final concentration, 10 mM) (see Table of Materials).
- Top up to 1 L with 1x Danieau solution (see Table of Materials). The solution must be kept in dark or wrapped with foil.
- On the day of the experiment, bring 500 mL of 2x AR solution to the zebrafish facility, and add 500 mL of system water (from the aquarium) to make 1x working AR solution.
- Transfer fish from their tanks into a 1x working AR staining solution and leave for 15-20 min. Wash off the excess stain on the fish with 15 min 'washing' in the system water.
- Perform live Calcein Green staining.
- Prepare a bottle of 2x Calcein Green staining solution by adding 45 mg of Calcein powder into 900 mL of 1x Danieau solution (see Table of Materials). Use a magnetic stirrer to allow the powder to dissolve completely.
- Adjust the pH to 8. This is the 2x stock Calcein Green solution. The solution can be stored in the dark/wrapped with foil for up to 1 month.
- On the day of the experiment, bring 500 mL of 2x Calcein Green solution to the zebrafish facility, and add 500 mL of system water to make 1x working Calcein Green solution. It is recommended to use freshly made Calcein staining solution.
- Transfer fish from their tanks into 1x working Calcein Green staining solution and leave for 1-2 h. Depending on the size of the fish, a longer staining time may be required.
- Wash off the excess stain on the fish by 15 min 'washing' in the system water.
NOTE: Image the Calcein Green stained fish as soon as possible as this stain will fade in a few hours.
3. Alkaline phosphatase (ALP) staining on scales post-harvest
- Prepare the ALP staining solution by mixing 100 mM Tris (pH 9.5 with HCl) with 100 mM NaCl and 50 mM MgCl2.
- Use commercially available NBT/BCIP stock solution (see Table of Materials).
- Transfer the scales into tubes containing deionized water.
- Rinse the scales in ALP staining solution for 5 min.
- Meanwhile, prepare a working staining solution by adding 200 µL of the NBT/BCIP solution to 10 mL of ALP staining buffer. This needs to be freshly prepared at the time of staining.
- Stain the scales with the working ALP staining solution for 15 min.
- Stop the reaction by rinsing the scales with deionized water.
4. Von KOSSA staining on scales post-harvest
- Prepare 5% (w/v) silver nitrate solution.
- Prepare 5% (w/v) sodium thiosulfate solution.
- Transfer harvested scales into tubes containing deionized water.
- Incubate the scales in the silver nitrate solution for 40 min under strong light.
NOTE: Wear PPE (personal protective equipment) as silver nitrate leaves a stain that is difficult to remove. - Wash the scales twice for 5 min with deionized water.
NOTE: Silver nitrate waste must be collected and disposed of according to local guidelines. - Incubate the scales in sodium thiosulfate for 5 min.
- Wash the scales twice for 5 min with deionized water.
5. Colorimetric TRAP staining
NOTE: For the detailed procedure, please refer to previously published work18.
- Prepare the staining solutions.
- Prepare 10 mg/mL Naphthol AS-MX phosphate solution.
- Weigh 5 mg Naphthol AS-MX phosphate (see Table of Materials).
- In the fume hood, prepare a tube with 0.5 mL of N, N'-dimethylformamide and dissolve 5 mg of Naphthol AS-MX phosphate in 0.5 mL of N, N'-dimethylformamide to make a stock solution of 10 mg/mL of Naphthol AS-MX phosphate (shake gently).
- Prepare 1.6 mM of Fast Red Violet LB stock solution in 50 mM sodium tartrate.
- Prepare 50 mL of 0.1 M sodium acetate at pH 5 (add 0.41015 g of sodium acetate in 50 mL of deionized water and adjust the pH to 5 with 100% acetic acid).
- Dissolve 0.575 g sodium L-tartrate dibasic dihydrate (see Table of Materials) in 50 mL of 0.1 M sodium acetate buffer (pH = 5).
- Add 30 mg of fast red violet LB salt.
- Prepare PBS-0.1Tx (dissolve 500 µL of Triton-X in 500 mL of PBS).
- Prepare PBS-0.1Tw (dissolve 500 µL of Tween-20 in 500 mL of PBS).
- Prepare 10 mg/mL Naphthol AS-MX phosphate solution.
- Prepare a working TRAP solution.
- Mix both stock solutions together in the fume hood (0.5 mL of 10 mg/mL Naphthol AS-MX phosphate solution and 50 mL of 1.6 mM Fast Red Violet LB stock solution in 50 mM sodium tartrate).
NOTE: The working solution can now be used outside the fume hood. It can be stored in the fridge (wrapped in foil) for up to a month. Store in separate containers from antibodies or other reagents sensitive to fixatives. Keep away from light as much as possible. - Optional: prepare bleaching solution (on post-fixed samples that were TRAP stained) in PBS and 0.1% Tween-20 (PBS-0.1%Tw).
- Prepare a final concentration mix of 0.5% potassium hydroxide (KOH) (from 10% stock solution (w/v)) and 3% hydrogen peroxide (H2O2) (33% stock solution, stored in the fridge).
- Mix both stock solutions together in the fume hood (0.5 mL of 10 mg/mL Naphthol AS-MX phosphate solution and 50 mL of 1.6 mM Fast Red Violet LB stock solution in 50 mM sodium tartrate).
- Perform sample fixation.
- Fix scales in 4% PFA in 1x PBS rocking or rotating for 40 min at room temperature (RT).
- Remove the 4% PFA and wash with PBS-0.1Tx at least 3 times for 5 min each wash.
- Directly move to TRAP staining (the best approach) or store in PBS-0.1 Tx overnight.
- Perform TRAP staining of the scales.
- Remove the PBS-0.1 Tx solution and add the colorimetric TRAP working solution (step 5.2.1) to cover the samples fully (i.e., 1 mL in a 1.5 mL microcentrifuge tube).
- Incubate for 1-2 h at RT, rocking in the dark.
- Remove the staining solution and wash at least 3 times with PBS-0.1 Tw for 5 min each wash.
- Post-fix the samples in 4% PFA for 30 min at RT, gently rocking in the dark.
- Remove the PFA solution by washing 3 times with PBS-0.1 Tw for 5 min each step.
NOTE: The samples can be stored (in storage solutions/mounted on microscope slides) at 4 °C in the dark indefinitely.
6. Mounting of stained scales
- Prepare the mounting medium by adding 2.4 g Mowiol 4-88 crystals to 6 g of glycerol (see Table of Materials). Any preferred mounting media can be used.
- Add 6 mL of deionized water and leave on a magnetic stirrer for an hour.
- Add 12 mL of 0.2 M Tris (pH 8.5) and incubate at approximately 53 ° C until the Mowiol crystals fully dissolve.
- Clarify the solution by centrifuging at 2000 x g for 20 min at room temperature.
- Transfer the solution to a storage container. The mounting medium can be kept in the freezer for about 12 months. The defrosted Mowiol solution is stable for up to 1 month.
- Mount the scales on a microscope slide in mounting the medium, cover them with a coverslip and view them under a microscope.
7. Ex vivo culture of scales
NOTE: This step is adapted from de Vrieze et al.12.
- Before harvesting the scales in a sterile environment, prepare the osteogenic mediumand the culture plate.
- Osteogenic Culture Medium (OCM): In a sterile centrifuge tube, make 15 mL OCM in standard cell culture medium (high glucose, no glutamine, no phenol red) at the final concentrations of 1% bovine calf serum, 1% (200 mM L-alanyl-L-glutamine dipeptide in 0.85% NaCl), 1% antibiotic/antimycotic solution (100x) solution, 4 mM CaCl2, 10 mM β-Glycerophosphate, 1 nM sodium pyruvate and 1 % Amphotericin B (optional) (see Table of Materials).
- Pour the OCM into a sterile reagent reservoir, and using a multichannel pipette, add 100 µL to each well of the 96-well plate.
- Seal the 96-well plate and store the plate. OCM can be stored in the refrigerator for a short period (overnight to 1 week) and kept at 28 °C before use.
- Perform scale harvesting and PBS washing.
- Harvest the scales using sterile tweezers/forceps and deposit them in a Petri dish containing a sterile PBS solution.
- Perform plate setup.
- Transfer each scale from the Petri dish to each well on the 96-well plate with 100 µL of OCM (warmed to 28 °C) using sterile forceps.
- Once all scales are transferred to the plate, bring the plate under a stereomicroscope. Using a fine, sterile pipette tip to gently move each scale to the bottom of the well and keep them away from the side of the wells to avoid light reflection during image acquisition.
NOTE: This step is required only if performing live imaging. - Carefully transfer the plate to the incubator or Live Imaging System (LIS, see Table of Materials) (set at 28 °C, 5% CO2), where the scales can be cultured for up to 7 days.
- Refresh OCM (optional).
- Warm the OCM that was stored in the fridge to 28 ˚C.
- Carefully transfer the 96-well plate to a sterile environment. Using a multichannel pipette, aspirate 50 µL from each well. For this, press the tips against the walls and do not dip them until the bottom to avoid aspirating/moving/removing the scales. Use new tips for different conditions.
- Pour the OCM into a sterile reagent reservoir, and using the multichannel pipette add 60 µL of OCM to each well.
- Place the plate under the stereomicroscope and check for misplaced scales (too close to the walls, floating, or in a vertical position within the media). Reposition them with a fine, sterile pipette tip if needed.
- Place the plate back in the incubator/LIS.
NOTE: Do not refresh the medium if the experiment involves circadian rhythm (CR), as serum shock can affect the biological clock. Note if studying CR notes that light conditions also reset the clock.
- Perform post-culture scale staining and imaging (optional).
- After 6 days of culture, fix them and direct them for appropriate staining such as von Kossa and ALP mentioned above.
- Alternatively, if the experimental fish have a fluorescent tag, put the plate into a live imaging system.
NOTE: If using a live imaging system, ensure that it is set to a temperature and CO2 level appropriate for the experiment. Typically, in this study, 28 °C and 5% CO2 were used. Select appropriate filters and imaging timepoints (here, an interval of 2 or 4 h, which is sufficient to follow osteoblast migration, was used) and low magnification objectives (i.e., 4x) to ensure even imaging of the entire wells. This is because the scales tend to move inside the wells, particularly during media changes.
Representative Results
Scale regeneration
Scale regeneration can be tracked with a standard fluorescent stereomicroscope by imaging the flank of the zebrafish. Figure 3A shows changes in sp7/osx expression of the scales during scale regeneration in a 4-month-old zebrafish. Timepoints for flank imaging shown in Figure 3A are ontogenetic (original scales, before harvesting), day 2, day 4, and day 10 post-harvest. We typically use osx (also known as osterix or sp7) transgenic lines (either as a GFP (Tg(Ola.sp7:NLS-eGFP)19 or mCherry Tg(osterix:mCherry-NTR)pd46)17 to track changes in the skeletal system as it labels bone-making osteoblasts. Early patterning of newly formed scales can be seen on 2 days of regeneration. This early pattern of scale regeneration is disrupted in some cases, especially in skeletal mutants. By tracking the progress of regeneration, one can analyze the capacity and rate of regeneration. On scale harvesting day, individual imaging of harvested scales can be performed after flank imaging using the same stereomicroscope, as shown in Figure 3B. Regenerated scales have much higher osx expression compared to Day 0 ontogenetic scales because osteoblasts are required for the new bone formation.
Ex vivo scale culture
Although one can study the de novo bone formation process by tracking the regeneration process on the flank of the fish, we can also use this model to study the repair and healing of skeletal injuries with ex vivo scale culture by making an injury on the scales with a scalpel. Using a live imaging system, the repair can be tracked in real-time. Figure 4 shows a representative result of an injury healing response on an ontogenetic scale in 3 days into a culture where the osteoblasts are labeled with osx: mCherry. The injury site shown by the insets at the beginning of the culture shows a clear gap between osteoblasts and scale circuli (Figure 4A). One can monitor the migration of osteoblasts toward the injury site with time-lapse imaging. After 3 days in culture, the width of the gap is reduced, and the expression of theosx can be seen between the gap and the newly formed scale circuli (Figure 4B). Additionally, the morphology in terms of the shape of the osteoblasts, is more circular at the beginning of the culture and after 3 days, is more elongated. This change in osteoblast appearance is likely due to being in culture and not in its natural environment (attached to the fish).
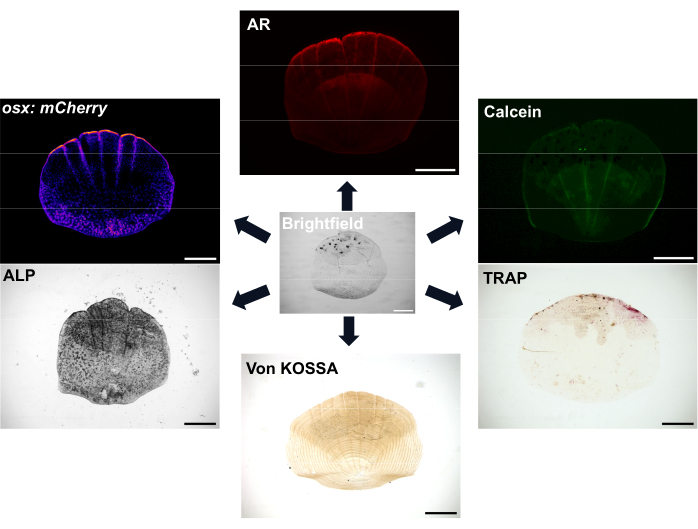
Figure 1: Examples of imaging options for scales. Osteoblasts can be visualized with osx/sp7 transgenic reporter lines (either in GFP or mCherry). ALP staining can be used to show the osteoblast activity. TRAP staining can show the activity of osteoclasts. Alizarin red (AR) and Calcein green are both dyes that can be used in live fish; AR labels mineralization and Calcein labels newly formed bone. The extent of mineralization can also be shown with von Kossa staining. Scale bars: 500 µm. Please click here to view a larger version of this figure.
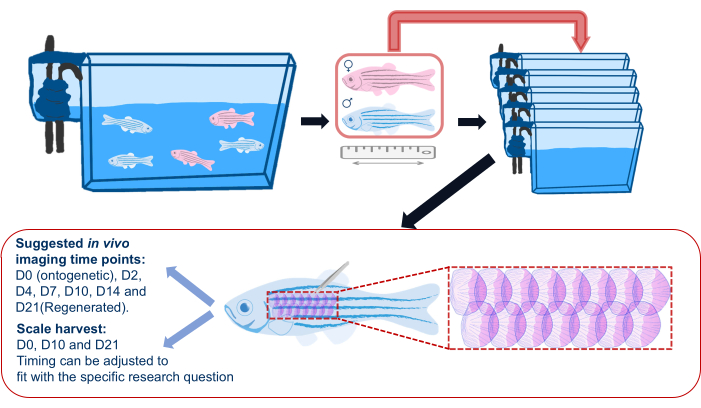
Figure 2: Schematic illustration of a scale regeneration experiment. Generic schematic of a scale regeneration experiment showing that fish is separated into individual tanks before the experiment. Length, sex, and health are recorded. The schematic also shows the suggested area on the flank of the fish to harvest scales and suggested imaging timepoints. Please click here to view a larger version of this figure.
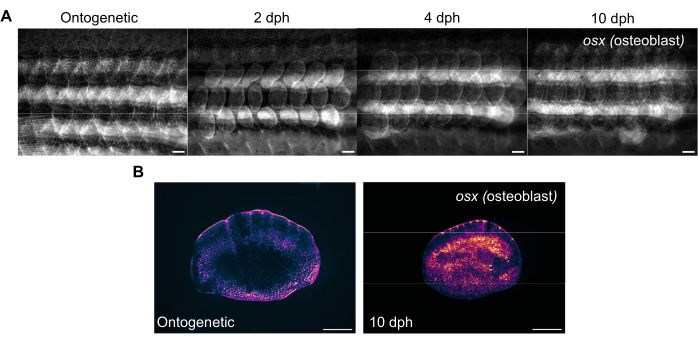
Figure 3: Representative images of osterix expression (to visualize osteoblasts) during scale regeneration taken from a 4-month-old zebrafish. (A) The flank images captured on day 0 (pre-harvest, ontogenetic scales), 2 dph (days post-harvest) day 2, 4 dph, and 10 dph for tracking changes in osx expression. Scale bars: 1 mm. (B) Representative images of an ontogenetic scale and a scale at 10 dph taken from the same fish as in panel (A). Note the increased levels of osx expression as shown in magenta in the center of the regenerating scales. Scale bars: 500 µm. Please click here to view a larger version of this figure.
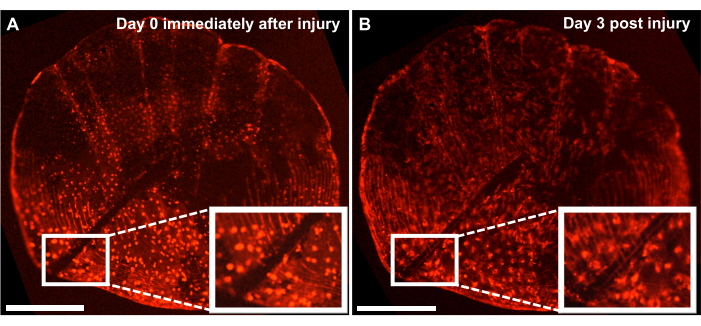
Figure 4: Representative results of skeletal injury repair response on an ontogenetic scale captured from a 4-month-old zebrafish using a live imaging system. (A) Ontogenetic scales with an injury made by scalpel on the same day as harvest (timepoint 0/day0) of the culture. (B) The same scale is shown 3 days after injury. Inset shows the region of injury. Scale bars: 500 µm. Please click here to view a larger version of this figure.
Discussion
The elasmoid scales of zebrafish, as a novel model for skeletal research, hold great potential to help our understanding of bone maintenance, regeneration, and injury repair. The abundance of scales on fish allows for medium to high-throughput compound screening while reducing the number of animals used and limiting intra-individual variation. Here, protocols for scale regeneration and ex vivo scale culture are presented to study regeneration and repair.
Some critical steps need to be considered when following this protocol. Careful removal of the scales is essential, especially when using a transgenic reporter line to limit the disturbance to the cell population caused by harvesting. If comparisons are to be made to ontogenetic scales, ensure that the region does not contain spontaneously regenerating scales (which can naturally occur through the lifespan of the fish). Ensure the environment and equipment are sterile for ex vivo culture to achieve optimal cell survival and minimal infection in culture.
Depending on the specific research question, adaptations can be made to the protocol, such as combining different transgenic reporter lines to visualize other cell types of gene expression profiles during regeneration and repair11,14.
The wide range of staining one can perform on the scales means that for each tested compound or condition one can look at its effects on the bone from different angles; while sp7/osx reporters can show osteoblast numbers, ALP staining can visualize osteoblast activity, TRAP staining can visualize osteoclast activity, Calcein green live staining can label newly forming bone and Alizarin red or von Kossa staining can show scale mineralization. Luciferase activity to quantify osteoblasts can also be used12. Combined with these staining techniques, one can learn the relative contribution of osteoblasts and osteoclasts to a given bone effect. Scales lack osteocytes, which are prevalent in mammalian bone and are the main drivers of bone mechanosensory response; scale repair and regeneration in this model are primarily driven by osteoblasts with subsequent remodelling by osteoclasts8,9. It is crucial to note that variation occurs between individuals and age groups20. To minimize this, the scale harvest area must be constant, as different locations can give rise to different scale morphologies, and fish from the same sibling groups are used so that age and size are consistent. However, because multiple scales can be harvested per fish, one can run more experiments using fewer fish, reducing intra-individual variability.
In summary, these protocols show experimental techniques that can be appliedto ontogenetic and regenerating scales. In conclusion, elasmoid scales show great potential as a skeletal model to aid the understanding of bone formation and repair; and will help to reduce animal use for high throughput osteoanabolic compound screening.
Divulgazioni
The authors have nothing to disclose.
Acknowledgements
We would like to thank Mathew Green of the Animal Service Unit for fish husbandry and Katy Jepson from the Wolfson Bioimaging Centre. CLH, DB, and QT were funded by Versus Arthritis (CLH Senior Fellowship 21937, DB and QT Intermediate Fellowship 22044), RR was funded by (NHMRC APP1158758). This work was also supported by the BBSRC grant (BB/T001984/1).
Materials
| 10x Phosphate buffered saline (PBS) | Gibco | 70013-016 | PBS |
| 12-Multichanel Pipette | Sartorius | 728230 | Multichanel pipette, Proline Plus Mechanical Pipette, 12 Channel, , 10-100 µL. |
| 15 mL Centrifuge tubes | Corning | 430791 | Centrifuge tube, CentriStar Cap, Polypropylene, RNAse/DNAse free, Non-pyrogenic |
| 4% Paraformaldehyde (PFA) | Sigma | P6148 | PFA |
| Alizarin red | Sigma | A5533 | |
| Amphotericin B | ThermoFisher Scientific | 15290026 | |
| Bemis Parafilm M Laboratory Wrapping Film | Fisher Scientific | 11772644 | Sealing film |
| Calcein powder | Sigma | C0875 | |
| Calcium Chloride | Thermo Scientific | L13191.30 | |
| Corning 96 well plate | Corning | 3596 | 96-well-plate, Clear, Flat Bottom Polystyrene TC-treated Microplates, Individually Wrapped, with Lid, Sterile |
| Cover slips | VWR | 631-0146 | |
| Cytiva HyClone Iron-Supplemented Calf Serum | Fisher Scientific | SH30072.03 | |
| Danieau | Sigma | ||
| DMEM | Life Technologies | 31053 | |
| Falcon tubes | Corning | 430828 | |
| Fast Red Violet LB stock solution | Sigma | F3381 | |
| GlutaMAX Supplement | Life Technologies | 35050 | |
| Glycerol | Sigma | 81381 | |
| Hepes | Sigma | H3375 | |
| Incubator | X | Incubator, Set up to 28 °C and 5% CO2 | |
| IncuCyte Zoom | Sartorious | X | Live Imaging System, Set up to 28 °C and 5% CO2 |
| Leica stereomicroscope | X | Sterioscope | |
| L-tartrate dibasic dihydrate | Sigma | 228729 | |
| Mgcl2 | BDH Laboratory Sup. | 261237T | |
| Microscope slides | Epredia | J2800AMNZ | |
| Mowiol 4-88 | Sigma | 9002-89-5 | |
| MQ water | X | ||
| N, N’-dimethylformamide (Merck: D4451) | Merck | D4451 | |
| NaCL | Fisher Chemical | S/3120/53 | |
| Naphthol AS-MX phosphate | Merck | N4875 | |
| NBT/BCIP solution | Sigma | #000000011681451001 | |
| Penicillin-Streptomycin | Life Technologies | 15140 | |
| Petri Dishes | Corning | 430589 | 35 mm sterile Petri dish, Non-treated, Nonpyrogenic, Polystyrene. |
| Reagent Reservoir | Startub | E2310-1025 | 25mL Reagent Reservoir |
| Silver nitrate | Sigma | 209139 | |
| Sodium acetate | Sigma | 52889 | |
| Sodium beta-glycerophosphate pentahydrate | Thermo Scientific | L03425.14 | |
| Sodium pyruvate solution | Sigma | S8636 | |
| Sodium tartrate | Sigma | S4797 | |
| Sodium thioculphate | Sigma | 563188 | |
| Tricaine methane sulfonate (MS222) | Sigma | E10521 | |
| Tris | Sigma | 252859 | |
| Triton-X100 | Sigma | T8787 | |
| Tween-20 | SLS | CHE3852 | |
| Tweezers Number 5 | Dumont | 500341 | Tweezer, INOX, biology grade |
| Zebrafish tanks | Tecniplast | ZB30BCP | 3.5 L – 28 cm x 11 cm x 17 cm |
| Zebrafish tanks | Tecniplast | ZB30BCP | 1 L – 28 cm x 7 cm x 11 cm |
Riferimenti
- Tobias, J. H., et al. Opportunities and challenges in functional genomics research in osteoporosis: report from a workshop held by the causes working group of the osteoporosis and bone research academy of the Royal Osteoporosis Society on October 5th 2020,". Frontiers in Endocrinology. 11, (2021).
- Busse, B., Galloway, J. L., Gray, R. S., Harris, M. P., Kwon, R. Y. Zebrafish: An emerging model for orthopedic research. Journal of Orthopaedic Research. 38 (5), 925-936 (2020).
- Dietrich, K., et al. Skeletal biology and disease modeling in zebrafish. Journal of Bone and Mineral Research. 36 (3), 436-458 (2021).
- McGowan, L. M., Kague, E., Vorster, A., Newham, E., Cross, S., Hammond, C. L. Wnt16 elicits a protective effect against fractures and supports bone repair in zebrafish. JBMR Plus. 5 (3), 10461 (2021).
- Sehring, I., Weidinger, G. Zebrafish fin: Complex molecular interactions and cellular mechanisms guiding regeneration. Cold Spring Harbor Perspectives in Biology. 14 (7), 040758 (2022).
- Bergen, D. J. M., Kague, E., Hammond, C. L. Zebrafish as an emerging model for osteoporosis: a primary testing platform for screening new osteo-active compounds. Frontiers in Endocrinology. 10 (6), (2019).
- Aman, A. J., Fulbright, A. N., Parichy, D. M. Wnt/β-catenin regulates an ancient signaling network during zebrafish scale development. Elife. 7, 37001 (2018).
- Iwasaki, M., Kuroda, J., Kawakami, K., Wada, H. Epidermal regulation of bone morphogenesis through the development and regeneration of osteoblasts in the zebrafish scale. Biologia dello sviluppo. 437 (2), 105-119 (2018).
- Metz, J. R., de Vrieze, E., Lock, E. J., Schulten, I. E., Flik, G. Elasmoid scales of fishes as model in biomedical bone research. Journal of Applied Ichthyology. 28 (3), 382-387 (2012).
- Cox, B. D., De Simone, A., Tornini, V. A., Singh, S. P., Di Talia, S., Poss, K. D. In toto imaging of dynamic osteoblast behaviors in regenerating skeletal bone. Current Biology. 28 (24), 3937-3947 (2018).
- Tonelli, F., et al. Zebrafish: A resourceful vertebrate model to investigate skeletal disorders,". Frontiers in Endocrinology. 11, (2020).
- de Vrieze, E., Zethof, J., Schulte-Merker, S., Flik, G., Metz, J. R. Identification of novel osteogenic compounds by an ex-vivo sp7: Luciferase zebrafish scale assay. Bone. 74, 106-113 (2015).
- De Vrieze, E., Moren, M., Metz, J. R., Flik, G., Lie, K. K. Arachidonic acid enhances turnover of the dermal skeleton: Studies on zebrafish scales. PLoS One. 9 (2), 89347 (2014).
- Bergen, D. J. M., et al. Regenerating zebrafish scales express a subset of evolutionary conserved genes involved in human skeletal disease. BMC Biology. 20 (1), 21 (2022).
- Bergen, D. J. M., et al. High bone mass disorders: New insights from connecting the clinic and the bench. Journal of Bone and Mineral Research. , (2022).
- Pasqualetti, S., Banfi, G., Mariotti, M. Osteoblast and osteoclast behavior in zebrafish cultured scales. Cell and Tissue Research. 350 (1), 69-75 (2012).
- Singh, S. P., Holdway, J. E., Poss, K. D. Regeneration of amputated zebrafish fin rays from de novo osteoblasts. Developmental Cell. 22 (4), 879-886 (2012).
- Ethiraj, L. P., Fong, E. L. S., Liu, R., Chan, M., Winkler, C., Carney, T. J. Colorimetric and fluorescent TRAP assays for visualising and quantifying fish osteoclast activity. European Journal of Histochemistry. 66 (2), 3369 (2022).
- DeLaurier, A., et al. Zebrafish sp7:EGFP: a transgenic for studying otic vesicle formation, skeletogenesis, and bone regeneration. Genesis. 48 (8), 505 (2010).
- Carnovali, M., Banfi, G., Mariotti, M. Age-dependent modulation of bone metabolism in zebrafish scales as new model of male osteoporosis in lower vertebrates. Geroscience. 43 (2), 927-940 (2021).
