Leptospirose hos mennesker stammer hovedsakelig fra miljøkilder 1,2. Tilstedeværelsen av Leptospira i innsjøer, elver og bekker er en indikator på leptospiroseoverføring blant dyreliv og husdyr og produksjonsdyr som til slutt kan komme i kontakt med disse vannmassene 1,3,4. Videre har Leptospira blitt identifisert i ikke-naturlige kilder, inkludert kloakk, stillestående og vann fra springen 5,6.
Leptospira er en verdensomspennende distribuert bakterie 7,8, og miljøets rolle i bevaring og overføring har blitt godt anerkjent. Leptospira kan overleve i drikkevann under variabel pH og mineraler9, og i naturlige vannmasser1. Den kan også overleve i lange perioder i destillert vann10, og under konstant pH (7,8) kan den overleve opptil 152 dager11. Videre kan Leptospira samhandle i bakterielle konsortier for å overleve tøffe forhold12,13. Det kan være en del av biofilm i ferskvann med Azospirillum og Sphingomonas og er til og med i stand til å vokse og tåle temperaturer over 49 ° C14,15. Det kan også formere seg i vannmettet jord og forbli levedyktig i opptil 379 dager16, og bevare sin evne til å forårsake sykdommen så lenge som et år17,18. Imidlertid er lite kjent om økologien i vannlegemer og hvordan den er fordelt i dem.
Siden oppdagelsen var studien av slekten Leptospira basert på serologiske tester. Det var ikke før det nåværende århundre at molekylære teknikker ble mer utbredt i studiet av denne spirochaete. Dot-blot har knapt blitt brukt til identifikasjon ved hjelp av (1) en isotopisk sonde basert på 16S rRNA og på en inter-enkel sekvensrepetisjon (ISSR) 19,20, (2) som en nanogoldbasert immunoassay for human leptospirose anvendt på urin21, eller (3) som en antistoffbasert analyse for storfe urinprøver22. Teknikken falt ut av bruk fordi den opprinnelig var basert på isotopiske prober. Det er imidlertid en velkjent teknikk som, kombinert med PCR, gir forbedrede resultater, og det anses som trygt på grunn av bruk av ikke-isotopiske prober. PCR spiller en avgjørende rolle i anrikningen av Leptospira DNA ved å amplifisere et spesifikt DNA-fragment som kan finnes i spormengder i en prøve. Under hver PCR-syklus dobles mengden av det målrettede DNA-fragmentet i reaksjonen. På slutten av reaksjonen har amplikonen blitt multiplisert med en faktor på mer enn en million23. Produktet forsterket av PCR, ofte ikke synlig i agaroseelektroforese, blir synlig gjennom spesifikk hybridisering med en DIG-merket sonde i dot-blot 24,25,26.
Dot-blot-teknikken er enkel, robust og egnet for mange prøver, noe som gjør den tilgjengelig for laboratorier med begrensede ressurser. Det har blitt brukt i en rekke bakteriestudier, inkludert (1) orale bakterier27, (2) andre prøvetyper som mat og avføring28, og (3) identifisering av unculturable bakterier29, ofte i samsvar med andre molekylære teknikker. Blant fordelene som tilbys av dot-blot-teknikken er: (1) Membranen har en høy bindingskapasitet, i stand til å binde over 200 μg / cm2 nukleinsyrer og opptil 400 μg / cm2; (2) Dot-blot-resultater kan tolkes visuelt uten å kreve spesialutstyr, og (3) de kan enkelt lagres i årevis ved romtemperatur (RT).
Slekten Leptospira har blitt klassifisert i patogene, mellomliggende og saprofytiske klader30,31. Skillet mellom disse kladene kan oppnås basert på spesifikke gener som lipL41, lipL32 og 16S rRNA. LipL32 er tilstede i de patogene kladene og utviser høy sensitivitet i ulike serologiske og molekylære verktøy, mens den er fraværende i saprofyttarter21. Housekeeping-genet lipL41 er kjent for sitt stabile uttrykk og brukes i molekylære teknikker32, mens 16S rRNA-genet brukes til klassifisering.
Denne metoden kan brukes på store mengder vann når de har blitt konsentrert ved sentrifugering. Det tillater vurdering av ulike punkter og dybder i en vannkropp for å oppdage tilstedeværelsen av leptospiralt DNA og kladen som den tilhører. Dette verktøyet er verdifullt for både økologiske og generelle screeningformål, og kan også brukes til å oppdage andre ikke-dyrkbare bakterier som kan være tilstede i vann.
I tillegg er PCR- og dot-blot-analyser teknisk og økonomisk rimelige for et bredt spekter av laboratorier, selv de som mangler sofistikert eller dyrt utstyr. Denne studien tar sikte på å anvende digoksigeninbasert dot-blot for identifisering av de tre Leptospira-klader i vannprøver samlet fra naturlige vannmasser.
Bakterielle stammer
Tolv Leptospira serovarer (Autumnalis, Bataviae, Bratislava, Canicola, Celledoni, Grippothyphosa, Hardjoprajitno, Icterohaemorrhagiae, Pomona, Pyrogenes, Tarassovi, og Wolffi) ble inkludert i denne studien. Disse serovarene er en del av samlingen ved Institutt for mikrobiologi og immunologi, Fakultet for veterinærmedisin og zooteknikk, National Autonomous University of Mexico, og de brukes for tiden i mikroagglutineringstesten (LAR).
Alle Leptospira serovarer ble dyrket i EMJH, og deres DNA ble ekstrahert ved hjelp av et kommersielt DNA-ekstraksjonssett (se materialtabell). En genomisk DNA-blanding av de tolv serovarene ble brukt som en positiv kontroll for den patogene kladen Leptospira . Som positiv kontroll av mellomkladen Leptospira ble genomisk DNA fra Leptospira fainei serovar Hurstbridge-stamme BUT6 inkludert, og som positiv kontroll for Leptospira-saprofyttkladen ble også genomisk DNA fra Leptospira biflexa serovar Patoc-stammen Patoc I, inkludert.
Negative kontroller besto av et tomt plasmid, DNA fra ikke-relaterte bakterier (Ureaplasma urealyticum, Staphylococcus aureus, Brucella abortus, Salmonella typhimurium, Shigella boydii, Klebsiella pneumoniae, Acinetobacter baumannii og Escherichia coli) og PCR-gradig vann, som fungerte som ikke-malkontroll.
Vannprøver
Tolv prøvetakingsprøver ble samlet inn ved hjelp av en stratifisert-tilfeldig prøvetakingsmetode fra Cuemanco Biological and Aquaculture Research Center (CIBAC) (19° 16′ 54″ N 99° 6′ 11″ W). Disse prøvene ble tatt på tre dybder: overfladisk, 10 og 30 cm (figur 1A, B). Vanninnsamlingsprosedyrene påvirket ingen truede eller beskyttede arter. Hver prøve ble samlet i et sterilt 15 ml mikrosentrifugerør. For å samle prøven ble hvert rør forsiktig nedsenket i vannet, fylt på den valgte dybden og deretter forseglet. Prøvene ble oppbevart ved 22 °C og straks transportert til laboratoriet for behandling.
Hver prøve ble konsentrert ved sentrifugering i sterile 1,5 ml mikrosentrifugerør ved 8000 x g i 20 minutter ved romtemperatur. Dette trinnet ble gjentatt til alle prøvene ble konsentrert i ett rør, som deretter ble brukt til DNA-ekstraksjon (figur 1C).
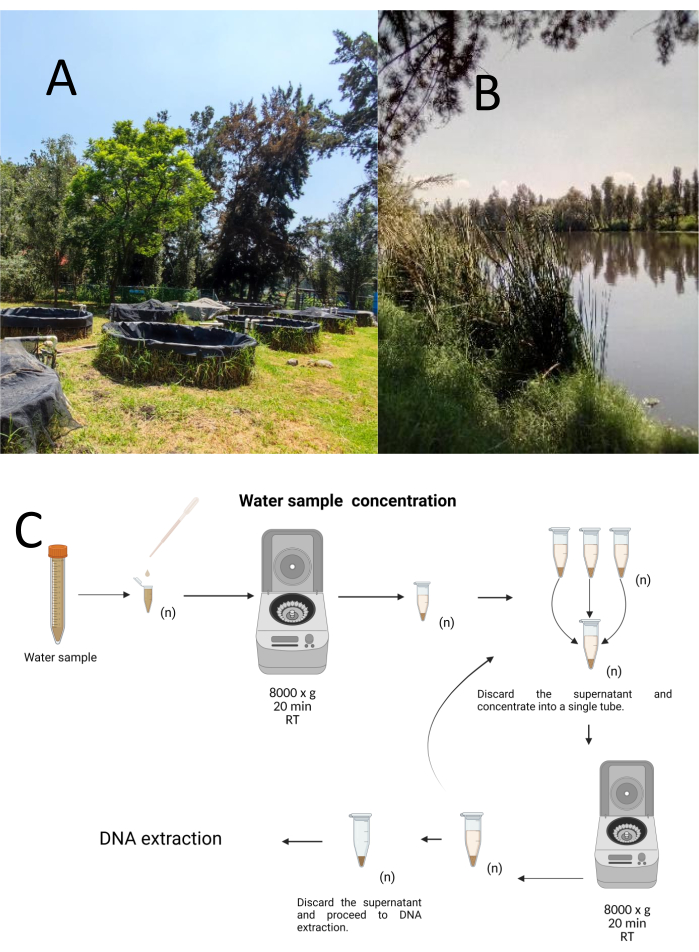
Figur 1: Konsentrasjon av vannprøver ved sentrifugering. (A) Vannprøvedammer, og (B) Naturlige bekker. (C) Sentrifugeringsbasert vannprøvebehandling i gjentatte trinn så mange ganger som nødvendig (n). Klikk her for å se en større versjon av denne figuren.
DNA-ekstraksjon
Totalt DNA ble isolert ved hjelp av et kommersielt genomisk DNA-sett i henhold til produsentens instruksjoner (se materialtabell). DNA-ekstraksjoner ble eluert i 20 μL elueringsbuffer, og DNA-konsentrasjon ble bestemt av et UV-spektrofotometer ved 260-280 nm, og lagret ved 4 °C til bruk.
PCR-amplifikasjon
PCR-målene var 16S rRNA-, lipL41- og lipL32-gener, som identifiserer DNA fra slekten Leptospira og tillater skillet mellom de tre kladene: patogen, saprofytisk og mellomliggende. Både primere og sondedesign var basert på tidligere arbeider av Ahmed et al., Azali et al., Bourhy et al., Weiss et al. og Branger et al.33,34,35,36,37. Sekvensen av hver sonde, primer og forsterkede fragment er beskrevet i tabell 1, og deres tilpasning til referansesekvenser er gitt i tilleggsfil 1, tilleggsfil 2, tilleggsfil 3, tilleggsfil 4 og tilleggsfil 5. PCR-reagensene og termosyklusforholdene er beskrevet i protokollavsnittet.
Forsterkningsprodukter ble visualisert ved elektroforetisk separasjon på en 1% agarosegel i TAE (40 mM Tris-base, 20 mM eddiksyre og 1 mM EDTA; pH 8,3), ved 60 V i 45 minutter med ethidiumbromiddeteksjon, som vist i tilleggsfigur 1. Genomisk DNA oppnådd fra hver serovar ble brukt med konsentrasjoner fra 6 x 106 til 1 x 104 genomiske ekvivalente kopier (GEq) i hver PCR-reaksjon, basert på genomstørrelsen til L. interrogans (4, 691, 184 bp) 38 for patogen Leptospira, genomstørrelsen til L. biflexa (3, 956, 088 bp) 39 for saprofytisk Leptospira, og genomstørrelsen til L. fainei serovar Hurstbridge-stamme BUT6 (4, 267, 324 bp) med tiltredelsesnummer AKWZ00000000.2.
Sensitiviteten til sondene ble vurdert med DNA fra hver patogen serovar, L. biflexa serovar Patoc stamme Patoc I og L. fainei serovar Hurstbridge stamme BUT6 i hvert eksperiment. For å vurdere spesifisiteten til PCR- og dot-blot-hybridiseringsanalysen ble DNA fra ikke-relaterte bakterier inkludert.
Tabell 1: PCR-primere og prober for å amplifisere produkter for å identifisere de patogene, saprofyte og mellomliggende klader av Leptospira. Klikk her for å laste ned denne tabellen.
Dot-blot hybridiseringsanalyse
Teknikken kalles dot-blot fordi hullene der DNA-prøven er plassert har en prikkform, og når de suges for å festes på plass ved vakuumsuging, får de denne formen. Denne teknikken ble utviklet av Kafatos et al.40. Teknikken tillater semi-kvantifisering av Leptospira i hver PCR-positiv prøve. Protokollen består av en denaturering med NaOH 0,4 M ved romtemperatur, prøver med Leptospira DNA fra 30 ng til 0,05 ng, tilsvarende 6 x 106 til 1 x 104 leptospirer, blir blottet på en nylonmembran med et 96-brønns dot-blot-apparat. Etter immobilisering er DNA bundet til membranen ved eksponering for 120 mJ UV-lys. Hver DNA-sonde er konjugert med digoksigenin-11 dUTP ved et terminalt transferasekatalysetrinn i 3′-enden (Digoxigenin er et plantesteroid oppnådd fra Digitalis purpurea, brukt som reporter41). Etter den strenge hybridiseringen av den merkede DNA-sonden (50 pmol) ved den spesifikke temperaturen på mål-DNA, visualiseres DNA-hybridene ved kjemiluminescensreaksjonen med det alkaliske fosfataseantistoffet mot digoksigenin kovalent konjugert med dets substrat CSPD. Luminescensen fanges opp ved eksponering for røntgenfilm (figur 2).
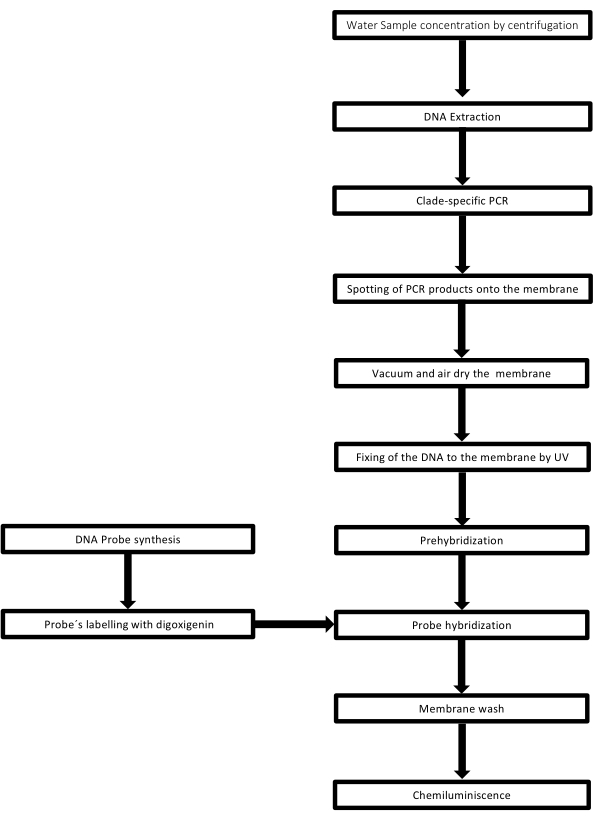
Figur 2: Trinn i prosedyren for PCR-dot-blot-analysen. Klikk her for å se en større versjon av denne figuren.
