غالبا ما يتطلب توضيح القواعد الجزيئية للوظائف الخلوية التعبير عن الحمض النووي المعدل وراثيا في زراعة الخلايا. للتعبير عنها ، يجب أن تخترق جينات التحوير الغشاء الانتقائي للخلية وتصل إلى النواة 1,2. لذلك ، فإن القدرة على تجاوز الحواجز المادية للخلية بشكل فعال والتلاعب بعملياتها المركزية هي ضرورة لتطبيق الجينات المحورة للكشف عن ظواهر بيولوجية جديدة. يستفيد أحد الأساليب من القدرة الجوهرية للفيروسات علىتوصيل الحمض النووي الأجنبي 3,4 والتعبير عنه.
الفيروس المرتبط بالغدي (AAV) هو واحد من أصغر فيروسات الثدييات: يحتوي جينوم الحمض النووي أحادي الشريط الذي يبلغ حجمه 4.7 كيلو قاعدة (kb) على جينين ، rep (للتكرار) وغطاء (للقفيصة) ، معبأ داخل قفيصة 60-mer icosahedral قياس 25 نانومتر. تحتوي جينات rep / cap على العديد من المروجين وإطارات القراءة ومنتجات لصق التي تشفر ما لا يقل عن تسعة بروتينات فريدة مطلوبة لتكاثر الفيروس وإنتاجه وتعبئته 5,6. بالإضافة إلى ذلك ، يحتوي كلا طرفي الجينوم على هياكل ثانوية تسمى التكرارات الطرفية المقلوبة (ITRs) الضرورية لتكرار الحمض النووي وتغليف الجينوم والمعالجة النهائية أثناء النقل7،8،9،10. ولوائح الاتصالات الدولية هي عناصر الحمض النووي الوحيدة اللازمة لتعبئة الجينوم في القفيصة، وبالتالي يمكن استنساخ AAV لأغراض توصيل الجينات المحورة عن طريق استبدال جينات الممثل/الغطاء الفيروسي باختيار الباحث للعناصر التنظيمية و/أو الجينات ذات الأهمية6. يستخدم AAV المؤتلف الناتج (rAAV) ، مع جينوم ناقل مهندس (VG) ، على نطاق واسع في عيادة العلاج الجيني البشري وقد حقق نجاحات11. استخدام الناقل لا يحظى بالتقدير الكافي في المختبر. يمكن ل rAAVs تحقيق التعبير الجيني المحورة بكفاءة في الخلايا المستزرعة لتلبية الاحتياجات التجريبية للباحث12.
الطريقة الأكثر شيوعا لإنتاج rAAV هي عن طريق نقل البلازميد الثلاثي إلى خلايا HEK293 أو 293T (الشكل 1). يحتوي البلازميد الأول، الذي يطلق عليه عادة بلازميد رابطة الدول المستقلة، على جين التحوير المطلوب المحاط بلوائح الاتصالات الدولية (pAAV). اعتمادا على التطبيق ، تتوفر بلازميدات رابطة الدول المستقلة ذات العناصر المشتركة ، مثل المروجين الأقوياء أو الأدوات القائمة على كريسبر ، للشراء. والثاني هو بلازميد pRep / Cap الذي يحتوي على جينات AAV rep و cap من النوع البري المقدمة في trans – أي على بلازميد منفصل غير ITR يحتوي على بلازميد يعبر عن العناصر التنظيمية والهيكلية التي تتفاعل بعد ذلك مع بلازميد رابطة الدول المستقلة – وبالتالي يسمى البلازميد المتحول. بالإضافة إلى إحاطة VG فعليا ، يؤثر القفيصة على الانتحاء الخلوي12,13. من خلال توفير جين الغطاء الخاص بالنمط المصلي في trans ، يمكن للباحثين بسهولة زيادة كفاءة النقل إلى أقصى حد عن طريق اختيار النمط المصلي القفيصة الأمثل للخلية المستهدفة المحددة. أخيرا ، باعتباره Dependoparvovirus ، يتطلب AAV فيروسا مساعدا لتنشيط تعبير rep / cap من المروجين الفيروسيين ، والذي تحققه الجينات المساعدة الفيروسية الغدية ، المقدمة على بلازميد ثالث مثل pAdΔF614,15. بعد 72 ساعة من انتقال البلازميد الثلاثي ، يمكن إطلاق الناقل من الخلايا المنتجة إلى وسط الاستزراع عن طريق دورات التجميد / الذوبان المتكررة. ثم يتم جمع محتويات اللوحة بأكملها ، ويتم إزالة الحطام الخلوي الكبير عن طريق الطرد المركزي ؛ طاف الوسائط الناتج هو إعداد rAAV خام جاهز لعمليات النقل النهائية.
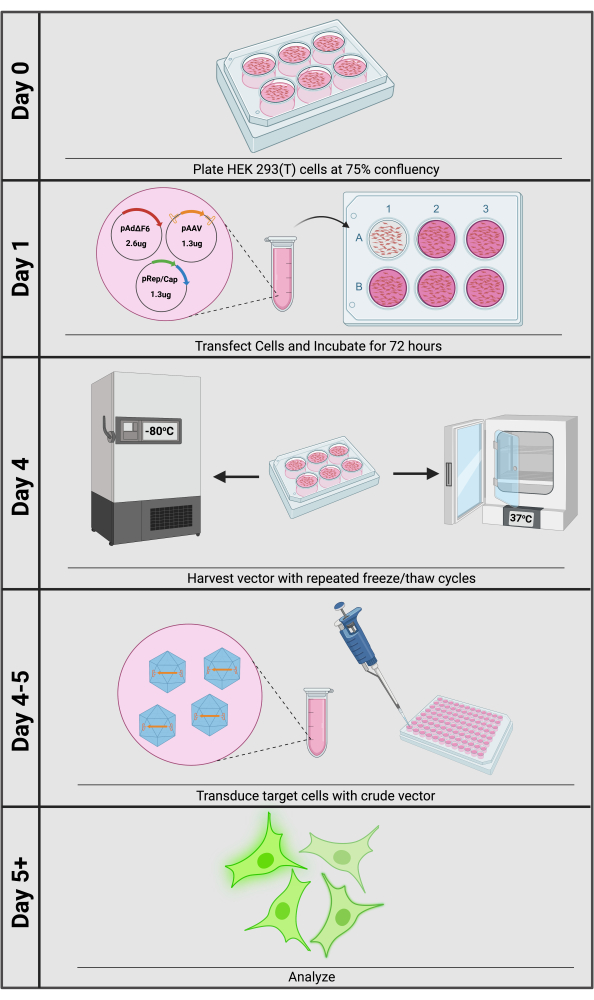
الشكل 1: نظرة عامة على إنتاج ناقلات rAAV الخام. يمكن إنجاز إنتاج ونقل الخام rAAV في غضون 5 أيام. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
قد يكون rAAV أكثر ملاءمة لتوصيل الجينات المحورة مقارنة بطرق النقل الأخرى ، والتي ترتبط عادة بالسمية الخلوية ، والكفاءة المنخفضة ، والكواشف والمعدات باهظة الثمن ، مثل التثقيب الكهربائي أو النقل الكيميائي / القائم على الدهون16,17. يتجاوز rAAV هذه العقبات وغالبا ما يوفر تعبيرا قويا عن الجينات المحورة بأقل قدر من السمية ، والحد الأدنى من الوقت العملي. الأهم من ذلك ، أن إنتاج rAAV وتطبيقه في زراعة الخلايا أمر بسيط ونادرا ما يتطلب تنقية الناقل من وسائط الاستزراع (الشكل 1). بالإضافة إلى ذلك ، لا يدمج rAAV VG الخاص به في جينوم المضيف ، على عكس توصيل الجينات المحورة Lentiviral ، وبالتالي يقلل من خطر حدوث طفرات إدراجية18. على الرغم من الفوائد المحتملة لاستخدام rAAV لتوصيل الجينات المحورة ، يجب مراعاة القيود. والأهم من ذلك، ينبغي ألا يتجاوز حجم جين التحوير، بما في ذلك لوائح الاتصالات الدولية، 4,9 كيلو بايت بسبب القيود المادية للقفيصة، مما يحد من قدرة الباحث على تقديم عناصر تنظيمية كبيرة وجينات محورة بشكل فعال. علاوة على ذلك ، نظرا لأن rAAV هو فيروس غير متكامل ، فإن النقل يؤدي إلى تعبير جين التحوير العابر في الخلايا المنقسمة وقد لا يكون عمليا للتعبير المستقر. ومع ذلك ، يمكن استخدام الطرق التي تستخدم قوالب Cas9 المزدوجة التي يتم تسليمها بواسطة rAAV والإصلاح الموجه بالتماثل (HDR) لإدراج تسلسلات ثابتة في مواقع جينومية محددة إذا رغب الباحثفي 19.
استنساخ
لا يقتصر بروتوكول الاستنساخ على بلازميد pAAV.CMV.Luc.IRES.EGFP.SV40 المستخدم أعلاه ويمكن تغييره بسهولة بناء على الاحتياجات التجريبية للباحث. ويتاح شراء العديد من البلازميدات المحتوية على لوائح الاتصالات الدولية بسهولة على الإنترنت. على سبيل المثال ، تتوفر البلازميدات التي تحتوي على كل من Cas9 وموقع استنساخ sgRNA ولكنها تتطلب خطوات إضافية قليلة مثل تلدين قليل النوكليوتيد ومعالجة PNK30. وبالإضافة إلى ذلك، يمكن العثور على البلازميدات التي تحتوي على موقع استنساخ متعدد (MCS) مع لوائح الاتصالات الدولية فقط وبدون عناصر تنظيمية داخلية31. إذا كان سيتم استخدام بلازميدات مختلفة ، فإن إنزيمات القطع (RE) المستخدمة في الهضم هي عادة العناصر الوحيدة التي قد تحتاج إلى تغيير في هذا البروتوكول. ومع ذلك ، فإن أحد قيود rAAV هو سعة الشحن المحدودة. ونظرا للقيود المادية للقفيصة، ينبغي ألا يتجاوز جينوم الناقل 4,9 كيلوبايت، بما في ذلك لوائح الاتصالات الدولية.
عند عزل البلازميد عن البكتيريا ، من الأهمية بمكان استخدام مجموعة midiprep أو maxiprep منخفضة السموم الداخلية أو خالية من الماكسيريب لتخفيف الضرر الذي يلحق بالخلايا أثناء نقل البلازميد الثلاثي أو التنبيث. غالبا ما تحتوي البلازميد من مجموعات miniprep على شوائب أعلى وتركيزات منخفضة وعدد أقل من الحمض النووي فائق الالتفاف ، وكلها يمكن أن تؤثر على الإنتاج النهائي ل rAAV وبالتالي لا ينصح به.
ومن الأهمية بمكان فهم هيكل لوائح الاتصالات الدولية وخصائصها أثناء الاستنساخ. أولا، من الصعب للغاية استخدام تفاعل البوليميراز المتسلسل من خلال لوائح الاتصالات الدولية. وينبغي تجنب تصاميم الاستنساخ التي تتطلب تضخيم تفاعل البوليميراز المتسلسل من خلال لوائح الاتصالات الدولية، بالإضافة إلى الحد من استخدام تقنية استنساخ تجميع جيبسون. وعلى هذا النحو، فإن استنساخ إنزيمات التقييد هو الطريقة المفضلة للاستنساخ في البلازميدات المحتوية على لوائح الاتصالات الدولية. وعلاوة على ذلك، قد لا تكون بعض البادئات لتسلسل سانجر متوافقة إذا كانت المنطقة المتسلسلة تحتوي على لوائح الاتصالات الدولية. وبدلا من ذلك، يوصى باستخدام الأعداد الأولية التي تتسلسل بعيدا عن لوائح الاتصالات الدولية وفي جسم الجينوم المتجه للحصول على نتائج تسلسل أكثر دقة. ثانيا، تكون لوائح الاتصالات الدولية عرضة للحذف وإعادة الترتيب والطفرات عند تحويلها إلى بكتيريا لتضخيم البلازميد32,33. للتخفيف من هذه الأحداث ، يوصى باستخدام سلالات بكتيرية مختصة تعاني من نقص إعادة التركيب ، مثل Stbl3 ، واحتضانها عند 30 درجة مئوية لإبطاء الانقسامات الخلوية. وأخيرا، لوحظ أن المستعمرات الأصغر قد تتوافق مع الحيوانات المستنسخة دون إعادة ترتيب أو حذف، لأن المستعمرات التي لا تحتوي على لوائح الاتصالات الدولية قد تمنح ميزة نمو وتكون أكبر. لذلك ، يوصى باختيار مستعمرات صغيرة.
إنتاج النواقل
يمكن أن يتأثر الإنتاج الناجح لناقل rAAV بعناصر متعددة. أحد العوامل الحاسمة هو صحة خلايا HEK293 أو 293T المستخدمة في النقل. بشكل عام ، تعتبر أرقام المرور المنخفضة مثالية ، حيث قد تظهر الخلايا ذات المرور العالي اختلافات في النمط الجيني والنمط الظاهري يمكن أن تقلل من عيار rAAV. بالإضافة إلى ذلك ، يجب أن تكون كثافة الخلايا المصنفة 75٪ -90٪ التقاء للإنتاج الفعال. تولد الخلايا المتناثرة غلات متجهة منخفضة نظرا لوجود عدد أقل من الخلايا المتاحة لإنتاج النواقل ، في حين أن الخلايا المتضخمة لن يتم نقلها بكفاءة.
تساهم الاختلافات بين الكثير من الكواشف ومخزون الخلايا والتباين العام من مختبر إلى مختبر في الاختلافات في كفاءة النقل وعيار الإنتاج. أحد العوامل القابلة للتحسين التي يمكن أن تؤدي إلى تحسينات العيار هو نسبة البلازميد: PEI في تفاعلات النقل. من الأهمية بمكان استخدام PEI MAX<. يوصى باستخدام نسبة البلازميد: PEI البالغة 1: 1 كنقطة انطلاق ، وإذا بدت كفاءة النقل أو التنبيغ ضعيفة ، فاختبر عدة نسب مختلفة. يكون تحسين العيار أسهل إذا كنت تستخدم جين التحوير مع قراءة مرئية ، مثل مراسل CMV.Luc.IRES.EGFP trangene المستخدم هنا كمواد أولية للاستنساخ. لإجراء التحسين ، اتبع خطوة البروتوكول 3 باستخدام لوحة 12 بئرا وتقليل كتل البلازميد وأحجام الكاشف بمقدار اثنين (كتلة البلازميد النهائية هي 2.6 ميكروغرام). اضبط حجم PEI وفقا لذلك ليتوافق مع النسب التي تتراوح من 1: 0.75 إلى 1: 3 ، مع زيادة الزيادات بمقدار 0.25 (الشكل 6). خفف كل تفاعل باستخدام 950 ميكرولتر من وسائط SF بعد 15 دقيقة. للراحة ، يمكن عمل مزيج رئيسي يحتوي على البلازميدات الثلاثية وسحبها بشكل فردي في أنابيب سعة 1.5 مل قبل إضافة PEI-see الملف التكميلي 2. حصاد المتجه ، وتحويل الخلايا ذات الأهمية ، والصورة. يتوافق البئر ذو أعلى كفاءة نقل (نسبة خلايا GFP +) مع أعلى عيار وأفضل نسبة من PEI: DNA.
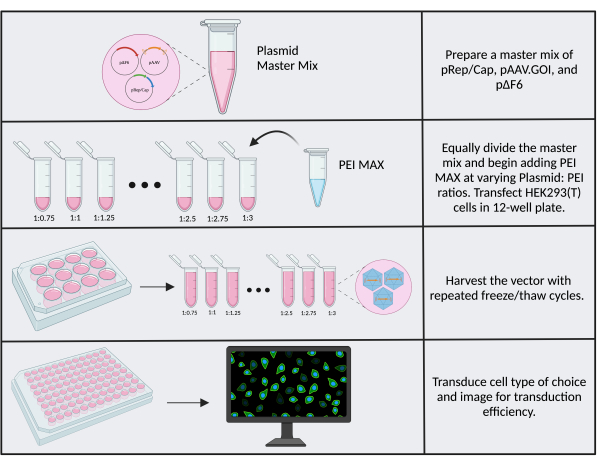
الشكل 6: سير عمل تحسين جزيرة الأمير إدوارد. رسم تخطيطي للخطوات المطلوبة لتحسين جزيرة الأمير إدوارد. نسب متعددة من البلازميد: يتم اختبار جزيرة الأمير إدوارد لتحديد النسبة المثلى. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
اعتبارات الحصاد والعيار
تعمل تقنية التجميد / الذوبان المستخدمة لحصاد ناقل rAAV على تحلل خلايا HEK293 بشكل فعال بطريقة متوافقة مع الاستخدام المباشر للمحللات الموضحة لتحويل الخلايا المستزرعة. يتم إطلاق بعض الأنماط المصلية rAAV ، مثل AAV1 و AAV8 و AAV9 من الخلايا أثناء إنتاج النواقل ويمكن حصادها من وسط الخلية المستزرع دون دورات التجميد / الذوبان34. عادة ما تنتج الطريقة الموصوفة هنا عيارا بترتيب 1 × 10 10 VG / mL عند استخدام قفيصة AAV2 ، و 1 ×10 11 VG / mL ل AAV8. في حين يمكن تحقيق عيارات أعلى عن طريق المنظفات أو غيرها من التحلل الكيميائي ، إلا أنها ضارة بالخلايا في الاستخدام النهائي وتتطلب تنقية rAAVs من المحلل. العيار السفلي هو أحد المقايضات التي يجب على الباحث مراعاتها عند تحديد ما إذا كانت المستحضرات الخام مناسبة لاحتياجاته البحثية ، ومع ذلك ، فإن التتر المنخفض الهامشي الناتج عن الطرق الموضحة هنا يمكن أن يحول العديد من أنواع الخلايا بشكل جيد للغاية (انظر النتائج التمثيلية). بالإضافة إلى كفاءة النقل وصحة الخلية ، يختلف عيار النواقل اعتمادا على القفيصة المستخدمة أثناء إنتاج rAAV وحجم وتسلسل جين التحوير داخل VG35.
عند حصاد مستحضرات النواقل الخام ، قد يكون الحمض النووي البلازميد الذي تم استخدامه أثناء نقل البلازميد الثلاثي موجودا ، وعلى الرغم من ندرته ، إلا أنه يؤدي إلى نقل المصب أثناء النقل. علاوة على ذلك ، قد ترتبط VGs غير المعبأة بالجزء الخارجي من القفيصات وتستدعي استجابة مناعية فطرية للحمض النووي العاري والأجنبي أحادي الشريط36,37. لذلك ، قد تتطلب أنواع الخلايا الحساسة هضم مستحضرات النواقل DNase وتنقيتها لإزالة VGs والبلازميد غير المعبأة.
إذا رغب المرء في حساب عيار المستحضر الخام ، فيمكن إجراء qPCR لتحديد عدد VG المعبأة داخل الجسيمات المقاومة للإنزيم DNase (DRP). باختصار ، يتم هضم كمية صغيرة من المستحضر الخام DNase لإزالة الحمض النووي البلازميد ، أو تلويث الأحماض النووية ، أو VG المعبأ جزئيا. ثم تخضع العينة ل qPCR ويتم تحديد كمية VG المحمية داخل DRPs ، مما ينتج عنه عيار بوحدات جينوم متجه لكل مل من التحضير الخام38. لا يوصى بإجراء معايرة متجهة باستخدام المقايسات المستندة إلى ELISA التي تحدد كمية التتر القفيصة. بالمقارنة مع فيروس AAV من النوع البري ، يعاني rAAV من نسبة من القفيصات الفارغة والمعبأة جزئيا39. ستقوم ELISA بتحديد كمية جميع القفيصات بغض النظر عن محتويات الجينوم الخاصة بها وستبالغ في تقدير وحدات الطاقة القابلة للتحويل الموجودة في المستحضر ، الأمر الذي يتطلب VG معبأ.
اعتبارات النقل
هناك العديد من العوامل التي تؤثر على عمليات نقل rAAV ويجب وضع اعتبارات مناسبة لأي تجربة جديدة. اعتمادا على المروج الذي يقود التعبير الجيني ، يمكن أن يحدث بداية التعبير في وقت مبكر من 4 ساعات بعد النقل (hpt) ، ويتم تحقيق ذروة التعبير عادة بمقدار 48 hpt. من المهم أن تضع في اعتبارك المدة الزمنية من البذر الأولي للخلايا إلى نقطة النهاية التجريبية. هذا لتقدير التقاء بداية الخلايا والتأكد من أنها لا تنمو بشكل زائد بنهاية التجربة. إذا أصبحت الخلايا مفرطة في الالتقاء ، فقد يتغير السلوك الخلوي بسبب استجابة الإجهاد ويمكن أن يربك النتائج التجريبية. يمكن لبعض أنواع الخلايا ، مثل U2-OS ، تحمل فرط النمو / تثبيط الاتصال بشكل جيد. بالإضافة إلى ذلك ، يمكنهم تحمل فترات طويلة (48 ساعة +) في وسط مكيف خال من المصل – وهو نتاج بروتوكول الإنتاج هذا. ومع ذلك ، قد تتطلب أنواع الخلايا الحساسة إضافة مصل أو تخفيف المستحضر الخام مع وسط نمو خاص للحفاظ على الصحة أثناء النقل. تعد كفاءة النقل المنخفضة قليلا من استخدام الوسائط المحتوية على المصل مقايضة محتملة لصحة الخلايا ويجب أن يأخذها الباحث في الاعتبار.
عادة ، بالنسبة للخلايا سريعة الانقسام ، يكون التقاء البداية بنسبة 50٪ تقريبا هو الأمثل للتطبيقات التي سيتم إنهاؤها 48 hpt. ومع ذلك ، يمكن تعديل التقاء وفقا لذلك بناء على احتياجات التجربة. لا ينصح بتحويل خطوط الخلايا الخالدة من النوع أحادي الطبقة التي تزيد عن التقاء 75٪ بسبب انخفاض كفاءة النقل. يتم تحويل معظم أنواع الخلايا المستزرعة بنجاح وصحية بعد الحضانة الليلية باستخدام مستحضرات rAAV الخام ، يليها تغيير إلى الوسائط الطازجة المحتوية على المصل في الصباح.
يعد النمط المصلي للقفيصة عاملا مهما يجب مراعاته عند إنتاج rAAV لتحويل خلية مستهدفة ، حيث أن القفيصة هي المحدد الأساسي للانتحاء الخلوي والتعبير الجيني المحوري اللاحق13. AAV2 هو نمط مصلي يستخدم على نطاق واسع نظرا لقدرته على تحويل العديد من أنواع الخلايا المستزرعة بشكل فعال12. يمكن أن تعزى خاصية AAV2 هذه إلى بروتيوغليكان كبريتات الهيبارين (HSPGs) الذي يعمل كعامل ارتباط أساسي ل AAV2 والمستويات العالية من HSPGs على الخلايا المستزرعة من التكيف إلى النمو في طبق40. القفيصات الأخرى ، مثل AAV9 ، أقل فعالية في تحويل أنواع الخلايا الواسعة ويمكن تفسيرها من خلال عوامل ارتباط الاعتماد التي لم يتم التعبير عنها في هذا الإعداد41. لذلك ، نوصي باستخدام AAV2 كخيار أول قفيصة في الخلايا المستزرعة إذا لم يتم اختبار الخلية المستهدفة المرغوبة مسبقا باستخدام rAAV في الأدبيات.
يرجى ملاحظة أن أحد القيود الرئيسية على مستحضرات النواقل الخام هو أنها غير مناسبة لتحويل النماذج الحيوانية. تتطلب الدراسات في الجسم الحي تنقية المستحضرات والخضوع لتقييم الجودة.
التعبير الجيني التحوير واعتبارات التكامل المحتملة
لا تؤدي rAAVs بشكل موثوق إلى التعبير الدائم عن جين التحوير. بمرور الوقت ، يمكن إسكات VGs وقد يتم إغلاق التعبير المعدل وراثيا بعد عدة مقاطع42. بالإضافة إلى ذلك ، تظل غالبية VGs عرضية ، ولا تحتوي rAAVs على بروتينات Rep الفيروسية التي من شأنها أن تتوسط في الاندماج المتكرر في جينوم المضيف كما هو الحال في العدوى الفيروسية من النوع البري أو تعزز تكرار VGs43. ونتيجة لذلك، ستضعف الحلقات في الخلايا المستحثة في النهاية بين الخلايا البنوية من خلال الانقسامات.
التكامل على المستوى القاعدي هو إمكانية لجميع مواد الحمض النووي المعدلة وراثيا التي يتم تسليمها. ومع ذلك ، فإن VGs المحتوية على ITR عرضة للتكامل بتردد أعلى44. لذلك ، يمكن ملاحظة التعبير الدائم عن جين التحوير في مجموعة فرعية صغيرة من الخلايا. يجب على المستخدمين التفكير في هذا الاحتمال خاصة عند استخدام rAAV لتوصيل إنزيمات قطع الحمض النووي ، مثل Cas9 ، لأن الفواصل المزدوجة التي تقطعت بها السبل قد تؤدي إلى تكرار أكبر للتكامل والتعبير الدائم45. في حين أن هذا يجعل rAAV مرشحا جيدا لتقديم قوالب إصلاح موجهة بالتماثل لوضع العلامات الداخلية أو إضافة الجينات ، يجب اعتبار إمكانية إدخال Cas919,46.
