Skin explant cultures
Feather bud development from ex vivo skin organ cultures can directly be observed under the microscope. Using the skin explant culture model of chicken stage 30 dorsal skin, the placodes are visible along the midline. The morphogenetic front then gradually propagates laterally toward the skin periphery with the formation of new feather primordia. These feather primordia will develop into short feather buds after 2 days in culture and long feather buds after 4 days in culture (Figure 1).
Skin recombination cultures
For skin recombination, when epithelia and mesenchyme are recombined, the original placodes disappear. New placodes will appear shortly after recombination and develop to form short feather buds and long feather buds after 2 and 4 days in culture, respectively. If the epithelium is rotated 90° relative to the mesenchyme, the orientation of the elongating buds will be determined by the epithelium (Figure 2).
Skin reconstitution cultures
For skin reconstitution, the ex vivo organ cultures appear homogeneous at first, and then dermal condensations with even spacing form simultaneously after 1 day in culture. It should be noted that the number of feather buds is dependent on the number of mesenchymal cells. Lower mesenchymal numbers induced fewer buds of a similar size to form11. Short feather buds will form after 2-3 days in culture and long feather buds will form after 4-5 days in culture (Figure 3).
Figure 4 shows summary diagrams of ex vivo skin organ culture, skin recombination organ culture, and skin reconstitution organ culture.
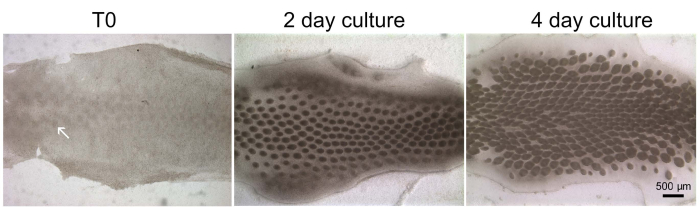
Figure 1: Ex vivo skin organ culture. Stage 32 chicken embryo skin is dissected in HBSS and cultured in 6-well culture inserts (T0, at time 0) for 2 and 4 days. The feather primordia develop into short feather buds after 2 days in culture and long feather buds after 4 days in culture. Dermal placodes are indicated by the white arrow. Note the buds in the midline are more mature than those on both lateral sides. This method has been modified from Jiang and Chuong4. Scale bar = 500 µm. Abbreviation: HBSS = Hank's buffered saline solution. Please click here to view a larger version of this figure.
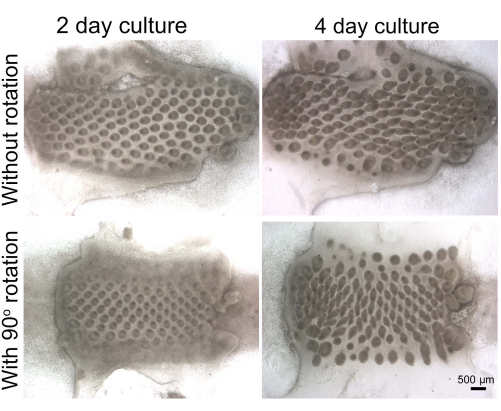
Figure 2: Ex vivo skin recombination organ culture. Stage 32 chicken embryo skin is dissected in HBSS, and epithelium and mesenchymal are separated in 2x CMF buffer. Skin epithelium and mesenchyme are recombined with or without rotation and cultured for 2 days and 4 days. The new placodes develop into short and long feather buds after 2- and 4-days in culture. If the epithelium is rotated 90° relative to the mesenchyme, the orientation of the new buds is determined by the epithelium12. This method has been modified from Chuong et al.11. Scale bar = 500 µm. Abbreviation: CMF = calcium-magnesium-free. Please click here to view a larger version of this figure.
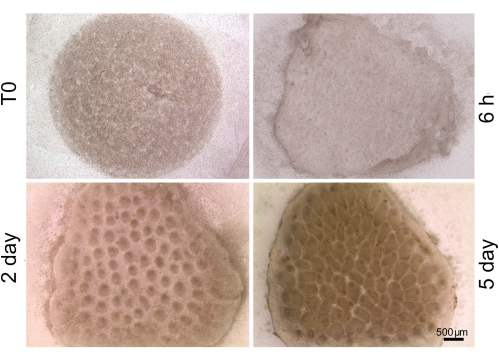
Figure 3: Ex vivo skin reconstitution organ culture. Stage 32 chicken embryo skin is dissected, and epithelium and mesenchyme are separated in 2x CMF buffer. The mesenchyme is dissociated into single cells by 0.1% collagenase and trypsin and pelleted at high cell density on a culture insert. The dermal cell pellet is reconstituted with an intact epithelium (T0, at time 0) and cultured for 6 h, 2 days, and 5 days. The explants appear homogeneous at first (6 h) and then dermal condensations with even spacing form simultaneously after 1 day in culture. Short feather buds will form after 2-3 days in culture and long feather bud form after 4-5 days in culture. This method has been modified from Jiang et al.13. Scale bar = 500 µm. Please click here to view a larger version of this figure.
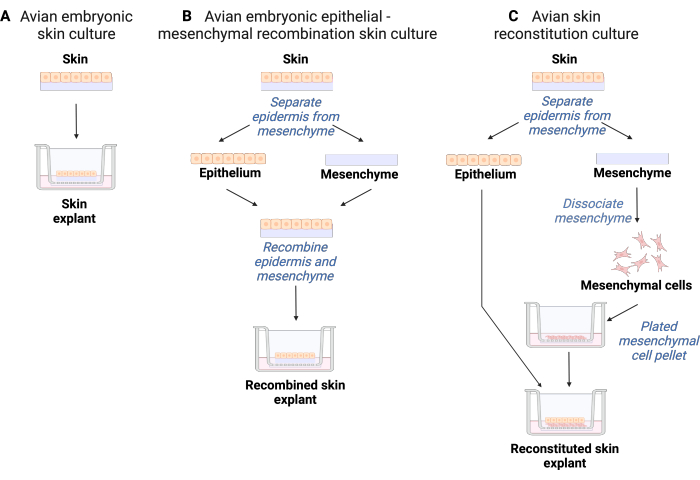
Figure 4: Diagrams of ex vivo skin organ culture models. The skin initiates at E6.5 as a single layer of epithelium overlaying mesenchymal cells. This is depicted at the top of the three explant methods. (A) Ex vivo skin organ culture. The skin is plated intact on top of a culture insert at the air: media interface at 37 °C in a 5% CO2 and 95% air incubator. (B) Ex vivo skin recombination organ culture. Skin epithelium is separated from the mesenchyme and then recombined before plating onto the cell culture insert at the air: media interface at 37 °C in a 5% CO2 and 95% air incubator. (C) Ex vivo skin reconstitution organ culture. Skin epithelium is separated from the mesenchyme. The mesenchyme is then dissociated into a single-cell suspension and the mesenchymal cells are pelleted in a centrifuge. The mesenchymal cells are then resuspended at a concentration of 2 × 107 cells/mL and 10 µL of the suspension placed onto the culture insert and then overlaid with an intact piece of epidermis. The culture is incubated at the air: media interface at 37 °C in a 5% CO2 and 95% air incubator. Please click here to view a larger version of this figure.
Supplemental Figure S1: Transduction of mesenchyme cells with GFP-expressing virus. Dissociated mesenchymal cells (2 × 107 cells/mL) were incubated for 3 h on ice with >107 infectious units/mL replication-competent avian sarcoma virus expressing Green Fluorescent Protein (GFP). The mesenchymal cells were then used to form reconstituted skin organ cultures as described in protocol section 3 above and photographed 24 h later. The data show that ~40% of mesenchymal cells were labeled with GFP. Please click here to download this File.
