Linsen er et klart og ovoid væv i øjets forreste kammer, der består af to celletyper, epitel- og fiberceller 1 (figur 1). Der er et monolag af epitelceller, der dækker linsens forreste halvkugle. Fiberceller er differentieret fra epitelceller og udgør hovedparten af linsen. De højt specialiserede fiberceller gennemgår en forlængelse, differentiering og modningsprogrammering, præget af forskellige ændringer i cellemembranmorfologi fra linseperiferien til linsecentret 2,3,4,5,6,7,8,9,10,11,12 , også kendt som linsekernen. Linsens funktion til finfokuslys, der kommer fra forskellige afstande på nethinden, afhænger af dets biomekaniske egenskaber, herunder stivhed og elasticitet 13,14,15,16,17,18,19. De komplekse interdigitationer af linsefibre er blevet antaget 20,21 og for nylig vist sig at være vigtige for linsestivhed 22,23.
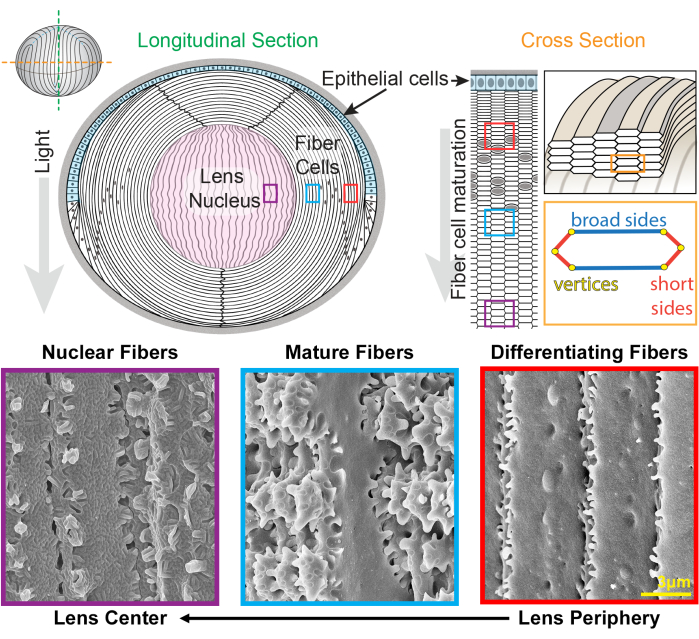
Figur 1: Linseanatomidiagrammer og repræsentative scanningelektronmikroskopibilleder (SEM) fra linsefibre. Tegneserien viser et langsgående (forreste til bageste fra top til bund) billede af det forreste monolag af epitelceller (skraveret i lyseblå) og en bulkmasse af linsefiberceller (hvid). Midten af linsen (skraveret i pink) er kendt som kernen og består af stærkt komprimerede fiberceller. Til højre afslører en tegneserie med tværsnit den aflange sekskantcelleform af linsefibre, der er pakket ind i et bikagemønster. Fiberceller har to brede sider og fire korte sider. Repræsentative SEM-billeder langs bunden viser de komplekse membraninterdigitationer mellem linsefiberceller i forskellige dybder af linsen. Fra højre har nydannede linsefibre i linseperiferien små fremspring langs de korte sider og kugler langs bredsiden (røde bokse). Under modning udvikler linsefibre store padledomæner, der er dekoreret med små fremspring langs de korte sider (blå kasser). Modne fiberceller besidder store padledomæner illustreret med små fremspring. Disse sammenkoblede domæner er vigtige for objektivets biomekaniske egenskaber. Fiberceller i linsekernen har færre små fremspring langs deres korte sider og har komplekse tunge-og-rille interdigitationer (lilla kasser). De brede sider af cellen viser en kugleformet membranmorfologi. Tegneserien blev ændret fra22,32 og ikke tegnet i skala. Skalabjælke = 3 μm. Klik her for at se en større version af denne figur.
Linsen vokser ved at tilføje skaller af nye fiberceller overlejret oven på tidligere generationer af fibre24,25. Fiberceller har en langstrakt, sekskantet tværsnitsform med to brede sider og fire korte sider. Disse celler strækker sig fra linsens forreste til bageste pol, og afhængigt af arten kan linsefibrene være flere millimeter lange. For at understøtte strukturen af disse aflange og tynde celler skaber specialiserede interdigitationer langs de brede og korte sider sammenlåsende strukturer for at opretholde linseformen og biomekaniske egenskaber. Ændringer i cellemembranform under fibercelledifferentiering og modning er blevet grundigt dokumenteret ved elektronmikroskopi (EM) undersøgelser 2,3,4,5,6,7,8,9,10,20,26,27,28,29 . Nydannede fiberceller har kugler og stikkontakter langs deres brede sider med meget små fremspring langs deres korte sider, mens modne fibre har sammenlåsende fremspring og padler langs deres korte sider. Nukleare fibre viser tunge-og-rille interdigitationer og kugleformet membranmorfologi. Lidt er kendt om de proteiner, der kræves til disse komplekse sammenlåsende membraner. Tidligere undersøgelser af proteinlokalisering i fiberceller har været afhængige af linsevævssektioner, som ikke tillader klar visualisering af den komplekse cellearkitektur.
Dette arbejde har skabt og perfektioneret en ny metode til at fiksere enkelt- og bundter af linsefiberceller for at bevare den komplekse morfologi og tillade immunfarvning for proteiner ved cellemembranen og i cytoplasmaet. Denne metode bevarer trofast cellemembranarkitektur, der kan sammenlignes med data fra EM-undersøgelser, og tillader farvning med primære antistoffer til specifikke proteiner. Vi har tidligere immunfarvede kortikale linsefibre, der gennemgår differentiering og modning22,23. I denne protokol er der også en ny metode til at plette fiberceller fra linsekernen. Denne protokol åbner døren til forståelse af mekanismerne for dannelse og ændringer i membraninterdigitationer under fibercellemodning og linsekernekomprimering.
