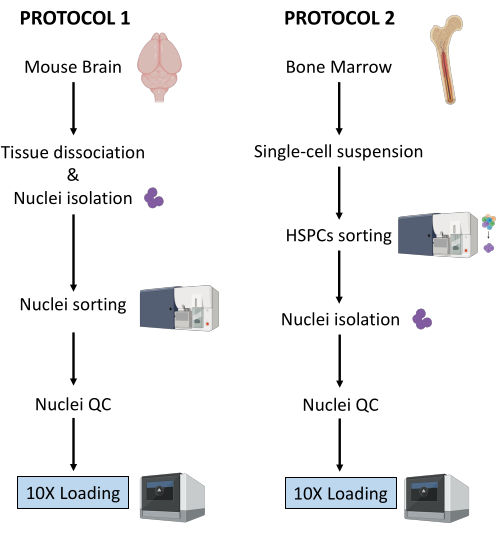
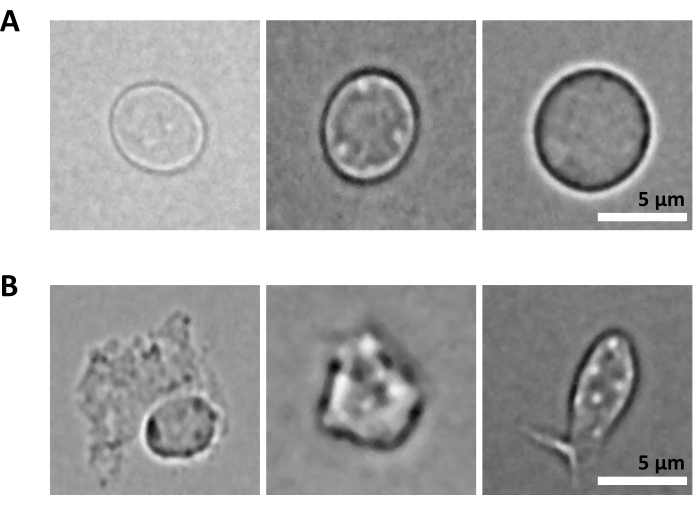
Forsøgsprocedurerne blev udført under streng overholdelse af protokoller godkendt af Komitéen for Etik i Dyreforsøg (CETEA). Til isolering af hjernekerner blev der anvendt 3 måneder gamle C57BL/6-mus. Til knoglemarvsisolering blev der anvendt 8 uger gamle C57BL/6J-hunmus, der vejede 18 g. 1. Oprensning af kerner fra musehjerne BEMÆRK: Brug altid latex- eller nitrilhandsker under proceduren. Det anbefales kraftigt at have to personer, der udfører eksperimentet, at få trin 1 til 3 (dvs. forberedelse af suspensionen med en enkelt kerne) udført af en person og trin 4 (dvs. forberedelse af sortereren) udført parallelt af en anden person. Da protokollen er meget tidsfølsom, er det afgørende at minimere prøvebehandlingstiden ved at have sortereren klar, så snart suspensionen med enkelte kerner er klar. Fremstilling af reagenser og materialerSteriliser forsigtigt dissektionsværktøjerne med autoklave (ved 121 °C i 20 min) og vask dem med ethanol 70% lige før brug. Tilbered en petriskål pr. prøve, fyldt med 2-3 ml iskold 1x Dulbeccos fosfatbufferede saltvand (DPBS). Mikrocentrifugen afkøles til 4 °C, fyldes en spand med is, og glasdouncehomogenisatoren lægges på is. Forbered kernelysebuffer ved at tilsætte digitonin til en slutkoncentration på 0,01%, 10 ml pr. Prøve. Forbered farvningsbufferen ved at tilsætte RNase-hæmmere til cellefarvningsbufferen for en slutkoncentration på 0,2 U / μL, 20 ml pr. Prøve. Forbered DPBS 0,04% BSA ved at tilsætte RNase-hæmmere til en slutkoncentration på 0,2 E / μL, 2 ml pr. Prøve. Forbered 1 ml fortyndet kernebuffer i henhold til Multiome-protokollen25. Opbevar alle reagenser og prøver på is. Dissektion af vævOfre mus ved hjælp af protokoller godkendt af institutionen. I denne protokol blev musene halshugget efter en overdosis ketamin/xylazin. Klip musehovedet med en saks og fjern hjernen fra kraniet som beskrevet i Meyerhoff et al.26. Overfør straks hjernen til en petriskål tilberedt med det iskolde 1x DPBS under et lysdiode (LED) -oplyst stereomikroskop. Skær hjernevævet med en skalpel for at adskille hjerneområder af interesse (f.eks. Entorhinal cortex, hippocampus, præfrontale cortex) og overfør hver region til en separat petriskål, der indeholder iskold 1x DPBS. Hold på is. Med en skalpel hakkes vævet i stykker på <0,5 cm for at lette homogeniseringen i det følgende trin. Med en P1000 mikropipette overføres hakket væv og 1x DPBS fra petriskålen til et 1,5 ml rør. Sørg for at bruge rør lavet af protein-lavbindende plast. Lad vævsstykkerne adskille sig ved tyngdekraften. Fjern forsigtigt overskydende 1x DPBS ved hjælp af en P1000 mikropipette.BEMÆRK: Efter dette trin er det muligt at snapfryse det hakkede væv ved at overføre proteinrørene med lav binding til tøris og derefter opbevare ved -80 °C, indtil der fortsættes med kerneisolering. Isolering af kernerFyld glasducen med 2 ml iskold kerner lysisbuffer med 0,01% digitonin. Tilsæt vævsstykkerne i dounce.BEMÆRK: Hvis du arbejder med friskfrosset væv, tilsættes hakket frosset væv direkte til kernernes lysbuffer 0,01% digitonin; Lad ikke vævet tø op før. Homogeniser ved hjælp af en glasdounce vævshomogenisator 25 gange med støder A og derefter 25 gange med støder B. Overfør homogenatet til et 15 ml rør. Tilsæt yderligere 2 ml iskold kerner lysebuffer med 0,01% digitonin og inkuber på is i 5 min. Centrifuger kerner ved 500 x g i 5 minutter ved 4 °C. Supernatanten fjernes med en mikropipette og tilsættes 4 ml iskolde kerner med 0,01% digitonin. Inkuber på is i 5 min og filtrer gennem en 40 μm cellesi. Centrifuger kernerne ved 500 x g i 5 minutter ved 4 °C, og supernatanten fjernes med en mikropipette. Der tilsættes 4 ml farvningsbuffer for at vaske kernerne og centrifugeres ved 500 x g i 5 minutter ved 4 °C. Supernatanten fjernes med en mikropipette og opslæmmes pelletten igen i 4 ml farvningsbuffer. Der filtreres gennem en 40 μm cellesi og centrifugeres ved 500 x g i 5 minutter ved 4 °C. Resuspender i 1 ml PBS med 0,04% BSA. Tæl kerner for at sikre konsistens af væv/kernepræparat på tværs af forskellige prøver. Det forventes at opnå lignende kernetal fra de samme hjerneområder:Tilsæt 10 μL 0,4% trypanblå til et tomt 0,5 ml rør. Der tilsættes 10 μL kerner og blandes 5x ved pipettering. Tæl kerner ved hjælp af en automatiseret celletæller efter leverandørens anbefalinger. Hold kernerne på is. Forbered kerner til sortering.BEMÆRK: De ekstraherede kerner indeholder 7-AAD, og denne farvning bruges til deres oprensning ved fluorescensaktiveret cellesortering (FACS).Overfør 100 μL kerner til et FACS-rør til den ufarvede kontrol. Der tilsættes 10 μL 7-AAD til de resterende kerner, og 5 minutter opbevares ved 4 °C. Sorter mindst 0,5 x 106 kerner efter FACS for at fjerne dubletter og snavs. Kernesortering ved hjælp af en FACSBEMÆRK: Mens kernesortering kan udføres på en lang række cellesorterere, er proceduren for brug af BD FACSAria Fusion- eller BD FACSAria III-instrumenterne beskrevet her. Det anbefales kraftigt, at kalibreringen og opsætningen af cellesorteringen udføres under tilsyn eller af en erfaren bruger af instrumentet. For at reducere prøvebehandlingstiden er det afgørende at have sortereren klar, så snart suspensionen med enkelte kerner er klar.Kalibrering af FACS-instrumentTænd cellesorteringen og computeren. Når softwaren er tilsluttet instrumentet, skal du starte den flydende opstartsprocedure. Vælg Cytometer > fluidisk opstart i hovedmenuen, og følg de fire trin. Klik på Udført efter at have gennemført hver. Indsæt 70 μm dysen, tænd for strømmen, og lad strømmen stabilisere sig i 15 minutter. Juster amplitude for at få dråbedannelse, og klik på Sweet Spot. Sæt filteret med neutral densitet (N.D) 1.0, og åbn cytometeropsætnings- og sporingsgrænsefladen (CST). Daglig kvalitetskontrol: Fortynd CST-perler i FACS-medium (se leverandørens anbefalinger) og udfør CST-kontrol. Når du er færdig, skal du udskifte N.D 1.0 med N.D 2.0. Fortynd Accudrops i FACS-medium (se leverandørens anbefalinger), og udfør faldforsinkelse som beskrevet i trin 6 til 10. Vælg eksperimentet Accudrop Drop Delay i eksperimentskabelonen, og åbn sorteringslayoutet for røret. Inde i det nederste kameravindue skal du klikke på Spænding og derefter på Optisk filter for at aktivere påføring af opladning på afbøjningspladerne og brug af et specifikt optisk filter foran kameraet. Sørg for, at kvadranten på højre side angiver 100. Juster om nødvendigt den røde laserskrue for at optimere laserpåvirkningen. Juster flowhastigheden for at nå hastigheden på 1.000 til 3.000 hændelser pr. Sekund. Klik på Sorter og annuller. Sørg for, at venstre kvadrant er lig med 100, og højre kvadrant er 0. Hvis venstre kvadrant er under 95, skal du udføre Automatisk forsinkelse. Klik på Spænding og derefter Test sortering. Kontroller kvaliteten af sidestrømmene i opsamlingsrørene. Hvis det er nødvendigt, kan du justere placeringen af sidestrømmene ved at flytte skyderne. Etablering af FACS-instrument til kernesortering.Start erhvervelsen af de ufarvede kerner. Disse bruges til at definere fremad- og sidespredningerne og detektorspændingen for 7-ADD-parameteren. Indstil parametrene, så 7-AAD-signalet for den ufarvede prøve falder inden for det første årti af logskalaen på prikplottet. Start med at erhverve røret med 7-AAD-farvede kerner, og definer kernepopulationerne ved hjælp af en gating-strategi baseret på (1) FSC-A/SCC-A og derefter FSC-H/SSC-H for størrelse og granularitet, (2) FSC-H/FSC-A for dubletterdiskrimination og (3) SSC-A/7-AAD for 7-AAD-positive kerner (se figur 2A). Sørg for, at strømmen og afbøjningen er stabile. I sidestrømskameraet skal du tænde testsorteringen, spændingen ON og bekræfte den nøjagtige faldsortering i et 1,5 ml rør monteret på venstre side. I vinduet Sorteringslayout skal du vælge den relevante population som defineret i trin 2 (ovenfor). I Målhændelser skal du vælge tærsklen i Kontinuerlig for at opnå mindst 0,5 x 106 kerner pr. prøve. Under Præcision skal du vælge 4-vejs renhed. Når du er klar, skal du klikke på Sorter og OK for at starte med kernesortering. Kvalitetskontrol og optælling af oprensede kernerBEMÆRK: Dette trin skal kun udføres under pilotforsøget til optimering af prøveforberedelsestrin med det formål at teste renheden af de sorterede kerner, der skal indlæses på 10X kromchippen. Når protokollen er fuldt optimeret, anbefales det ikke at udføre dette kvalitetskontroltrin i opfølgningseksperimenterne for at undgå unødvendigt spild af indsamlede kerner, der kan være tilgængelige i lavt antal.Renhedskontrol ved flowcytometriOverfør 10 μL af de sorterede kerner til et nyt FACS-rør indeholdende 90 μL DPBS med 2% varmeinaktiveret føtalt bovint serum (HI-FBS). Hent og registrer data efter sortering for at kontrollere sorteringens renhed og levedygtighed. Sørg for, at mindst 98 % af kernerne er placeret i interesseporten som defineret i 4.2 (se figur 2B). Tælling af de rensede kernerDe sorterede kerner centrifugeres i 5 minutter ved 500 x g og ved 4 °C, og supernatanten fjernes forsigtigt fuldstændigt ved hjælp af en mikropipette. Resuspender i 100 μL fortyndet kernebuffer. Tilsæt 10 μL 0,4% trypanblå til et tomt 0,5 ml rør. Der tilsættes 10 μL af de sorterede kerner og blandes 5x ved pipettering. Tæl kerner ved hjælp af en automatiseret celletæller efter leverandørens anbefalinger. Kernerne justeres til 3,5 x 106/ml, dvs. 16.000 kerner pr. 5 μL. Kvalitetskontrol af oprensede kerner ved mikroskopiBEMÆRK: Dette trin skal kun udføres under pilotforsøget til optimering af prøveforberedelsestrin for at teste kvaliteten af kernerne, der skal indlæses på 10X kromchippen. Når protokollen er fuldt optimeret, anbefales det ikke at udføre dette kvalitetskontroltrin i opfølgningseksperimenterne for at undgå unødvendigt spild af indsamlede kerner, der kan være tilgængelige i lavt antal.Sørg for, at mikroskopglas og dæksler er rene og støvfrie. Hvis det er nødvendigt, vask og skyl dækslerne med absolut ethanol og tør dem med fnugfri servietter. Der fordeles 25 μL poly-l-lysin i de glidehuller, der skal anvendes, og inkuberes i 10 minutter ved stuetemperatur (RT), beskyttet mod støv. Det overskydende poly-l-lysin fjernes, og der tilsættes 10 μL af den oprensede kernesuspension. Inkuber i 5 minutter ved RT, beskyttet mod støv. Tilsæt en dråbe monteringsmedium til hver brønd, undgå bobler. Placer en dækseddel oven på de frøede brønde. Dæk med papirservietter, og tryk dækslet fast for at fjerne det overskydende monteringsmedium. Pas på ikke at flytte dækslet, og rengør ikke det overskydende monteringsmedium. Tag flere billeder med et omvendt mikroskop med brightfield-lys og en minimumsforstørrelse på 40x. Udfør multiome-analyse.Fortsæt straks til Chromium Next GEM Single Cell Multiome ATAC + Gene Expression User Guide (CG000338 – Rev F)25. 2. Oprensning af kerner fra museknoglemarv, hæmatopoietiske stamceller og stamceller (HSPC’er) BEMÆRK Denne protokol beskriver oprensning af kerner fra tre undergrupper af knoglemarven HSPC’er: afstamning-c-Kit+Sca-1+ hæmatopoietiske stamceller (HSC), afstamning-c-Kit+Sca-1-CD34+FcγR- almindelige myeloide stamfædre (CMP) og afstamning- c-Kit+Sca-1-CD34+FcγR+ granulocyt-monocyt forfædre (GMP). Brug altid latex- eller nitrilhandsker under proceduren. Denne protokol er en tilpasning af 10X Genomics Proven Protocol – Nuclei Isolation for Single Cell Multiome ATAC + GEX sequencing (CG000365 – Rev C)27. Ændringer er blevet introduceret i den oprindelige protokol for at maksimere kernegendannelse. Det anbefales kraftigt at have to personer, der udfører eksperimentet, for at have trin 1. til 3. (dvs. fremstilling af enkeltcelleopløsningen) udført af en person, og trin 4 (dvs. forberedelse af sorteringen) udført parallelt af en anden person. Da protokollen er meget tidsfølsom, er det afgørende at minimere prøvebehandlingstiden ved at have sortereren klar, så snart enkeltcellesuspensionen er klar. Fremstilling af reagenser og materialerFyld en spand med is. Forbered FACS-bufferen: DPBS med 2% HI-FBS-opløsning (ca. 500 ml til 6 prøver) og filtrer gennem et 0,2 μm filter. Forbered opsamlingsmediet: DPBS med 10% HI-FBS-opløsning (500 μL pr. prøve), og filtrer gennem et 0,2 μm filter. Isolering af knoglemarvscellerOfre mus ved hjælp af protokoller godkendt af institutionen. I dette eksperiment blev musene ofret af cervikal dislokation efter en ketamin / xylazin overdosis. Sprøjt musenes mave og bagben med 70% ethanol. Brug sterile tang og saks til at lave et lille snit midt i underlivet og åbn bughinden fra bunden af bagbenene til mellemgulvet (supplerende figur 1). Lav et ekstra snit for hvert bagben vinkelret på det åbne bughinde, tag derefter fat i hver side af et af disse ekstra snit og træk det fra hinanden for at skrælle huden fra begge bagben forbi ankelleddet for at udsætte musklerne i begge bagben (supplerende figur 1A). Linj saksen langs rygsøjlen ved hofteleddet på det ene bagben for at skære benet ud uden at skære gennem lårbenet (supplerende figur 1B, C). Gentag det samme for det andet ben. For at isolere lårbenet skal du skære det meste af muskelvævet ud og derefter holde lårbenet og skinnebenet i hver hånd med fingerspidserne ved leddet (supplerende figur 1D, E). Fold forsigtigt benet mod den naturlige bøjning for at forskyde skinnebenet fra lårbenet (supplerende figur 1E), og klip derefter forsigtigt bindevævet med en saks for at adskille lårbenet og skinnebenet. Brug saksen med lette vridningsbevægelser til at forskyde rygraden fra lårbenets øverste ende (supplerende figur 1E). Rens det isolerede lårben med silkepapir for at fjerne den resterende muskel og bindevæv. Hold kold på is i en 12-brønds pladebrønd fyldt med 2 ml DMEM (1x) + GlutaMAX-I. Når alle lårbenene er samlet, skal du sikre dig, at muskel- og fibrøst væv fjernes fuldstændigt fra knoglen. Skær ikke knoglen åben for at (a) holde marven inde steril og (b) undgå at miste celler i brønden. Brug følgende trin til at skylle cellerne fra to lårben i en mus, tilpasset fra Haag og Murthy28. Forbered et 1,5 ml og et 0,5 ml rør. Tilsæt 150 μL af FACS-bufferen til 1,5 ml røret, stik derefter et hul i bunden af 0,5 ml røret ved hjælp af en 18 G nål og sæt 0,5 ml røret ind i 1,5 ml røret. Åbn den distale del af hvert lårben ved hjælp af musekirurgisk saks (supplerende figur 1F): Lås den distale epifyse mellem knivene, og tryk forsigtigt, mens du vender saksen for jævnt at løsne den distale epifyse uden at skære knoglen hårdt op. Hvis det lykkes, skal 4 fremspring være synlige i den nu eksponerede fyseende (supplerende figur 1G). De to lårben anbringes med den åbne ende nedad i det forberedte 0,5 ml rør, der er anbragt i et 1,5 ml rør indeholdende FACS-buffer (supplerende figur 1H). Anbring en 70 μm cellesi på et 50 ml rør og forvåd sien med 2 ml FACS-buffer. For at skylle knoglemarven centrifugeres rørene (hætterne åbnes) ved 12.000 x g, indtil centrifugen når 12.000 x g-værdien, og stop derefter straks centrifugen. Kontroller, at knoglemarvscellerne er pelleteret i 1,5 ml røret, og at lårbenene er hvide (før celleskylning er de røde) (supplerende figur 1I). Kassér 0,5 ml rørene med de 2 lårben. 150 μL supernatanten kasseres med en pipette. Resuspender pellet med en mikropipette i 1 ml ammoniumchlorid-kalium (ACK) lyserende buffer i 1-2 minutter ved RT for at lyse røde blodlegemer. Undgå længere inkubationstider, da de kan resultere i nedsat levedygtighed af kerneceller. Overfør til 50 ml røret gennem den forvædede 70 μm cellesi. Der tilsættes 10 ml FACS-buffer for at fortynde ACK-lyseringsbufferen, og hermed standses lysisen. Der centrifugeres ved 400 x g i 5 minutter ved 4 °C. Resuspender i 10 ml FACS-buffer ved først at resuspendere i 1 ml og derefter fylde op med 9 ml. Cellerne forberedes til tælling som beskrevet i punkt 1.3.8. Tæl cellerne ved hjælp af en automatiseret celletæller efter leverandørens anbefalinger. Det forventes at indsamle ca. 40 millioner celler fra 2 lårben. Farvning af knoglemarv HSPCCentrifuger cellerne ved 400 x g i 5 minutter ved 4 °C og resuspender pellet med en mikropipette i FACS-buffer til en slutkoncentration på 1 x 107 celler/ml. Med en P1000 mikropipette overføres suspensionen til et FACS-rør og filtreres gennem et 35 μm cellesidæksel. Forbered enkeltpletprøver reagensglas for hvert antistof, der er anført i tabel 1 , for at oprette kompensationer for fluorkromer på cellesortereren:Forbered et FACS-rør pr. antistof, og fyld glassene med 200 μL PBS. Der tilsættes 15 μL fluorkromkompensationsperler i hvert FACS-rør af fluorkromkonjugeret antistof. I FACS-rørene til ubejdsede og levende / døde enkeltfarvede celler tilsættes 500.000 celler i stedet for perler. Der tilsættes 1 μL af hvert fluorkromkonjugeret antistof (se tabel 1) i det tilsvarende FACS-rør. Tilsæt 0,5 μL levende/død plet i levende/død enkeltplet FACS-rør. Opbevares på is i 15 min beskyttet mod lys. Forbered blanding 1 og 2 som angivet i tabel 2.BEMÆRK: De antistofvolumener, der er angivet i tabel 2 , gælder for de antistoffer, der henvises til i materialetabellen. De skal optimeres til enhver ny antistofreference eller et andet parti af den samme antistofreference. Tilsæt 300 μL Mix 1 i prøveglasset, resuspender og opbevar i 15 minutter på is beskyttet mod lys. Tilsæt 300 μL Mix 2 i prøveglasset, resuspender og opbevar i 20 minutter på is beskyttet mod lys. Der tilsættes 3 ml FACS-buffer til de enkeltfarvede rør og de blandingsfarvede prøveglas. Spin ned ved 400 x g i 5 minutter ved 4 °C. Supernatanten kasseres forsigtigt med en mikropipette og pellettet resuspenderes i 500 μl FACS-buffer. Forbered et 1,5 ml rør fyldt med 500 μL opsamlingsmedium.BEMÆRK: Mix 1 fremstilles i DPBS, da det indeholder den levende/døde plet, der er væsentligt påvirket af HI-FBS. Når cellerne er farvet af Live/Dead, tilsættes Mix 2, som indeholder de fluorkromkonjugerede antistoffer resuspenderet i FACS-buffer indeholdende HI-FBS. Den eneste undtagelse er anti-CD16/32-antistoffet, der er inkluderet i antistofblanding 1 for at tjene som Fc-receptorblokker, der forhindrer den ikke-specifikke binding af de andre antistoffer, der tilsættes i det følgende trin. Cellesortering ved hjælp af en FACSBEMÆRK: Mens cellesortering kan udføres på en lang række cellesorterere, beskrives proceduren for brug af instrumenterne BD FACSAria Fusion eller BD FACSAria III her. Det anbefales kraftigt, at kalibreringen og opsætningen af cellesorteringen udføres under tilsyn eller af en erfaren bruger af instrumentet.Kalibrering af FACS-instrument: Se protokol 1, trin 4.1. Opsætning af FACS-instrument til cellesortering:Start erhvervelsen af de ufarvede celler. Disse bruges til at definere fremad- og sidespredningerne og detektorspændingen for hver fluorofor. Indstil parametrene, så fluorescerende signal for hver fluorofor falder inden for det første årti af logskalaen på prikplottet. Anskaf enkeltfarvekontroller for at konfigurere kompensationer manuelt (medianen af positive og negative populationer skal justeres), eller brug den automatiske beregningssoftware (hældningsmålinger). Sørg for, at kompensationskontrollerne matcher de eksperimentelle fluorkromer og detektorindstillinger. Optag 10.000 hændelser for celler og 5.000 hændelser for perler. Brug prøverøret (dvs. flerfarvede celler) til at definere cellepopulationer af interesse ved hjælp af gating-strategien vist i figur 3A. Følg trin 4-6 (nedenfor). For at identificere de tre knoglemarvs-HSPC’er af interesse (HSC, CMP og GMP) skal du starte gatingen ved at bruge størrelsen (FSC-A) og granulariteten (SSC-A) til at gate på leukocytter og derefter FSC-H / FSC-A for at diskriminere dubletter. Baseret på SSC-A / død cellemarkør, gate levende celler. Brug Afstamning/c-Kit til at vælge celler, der er afstamningsnegative og udtrykker mellemliggende til høje niveauer af c-Kit. Gennem c-Kit/Sca-1, gate på lineage-c-Kit+ Sca-1+ (LKS+) HSC’er, en af de tre populationer af interesse. Blandt de myeloide forfædre (afstamning-c-Kit+Sca-1-) skal du bruge FcγR/CD34 til at udelukke CD34-FcγR- megakaryocyt- og erythroid-stamfædre (MEP), mens CD34+ FcγR-CMP samt CD34+FcγR+ GMP inkluderes i de cellepopulationer, der skal sorteres. Sørg for, at strømmen og afbøjningen er stabil. I sidestrømskameraet skal du tænde testsorteringen, spændingen TIL og bekræfte den nøjagtige faldsortering i et 1,5 ml rør monteret på venstre side. I vinduet Sorteringslayout skal du vælge de(n) relevante population(er) (dvs. “LKS+” og “CD34+ myeloide forfædre” vist i dette eksempel). Under Enhed skal du vælge 2 Tube. Under Præcision skal du vælge Renhed. I Målhændelser skal du vælge Kontinuerlig for at sortere mellem 160.000 og 200.000 LKS+ og CD34+ myeloide forfædre. Der tilsættes 500 μL FACS-buffer til cellesuspensionen, og 1 ml af prøven overføres ved filtrering i et nyt FACS-rør med 35 μm-celle sikappe for at sikre, at alle celler er i en enkelt suspension lige før erhvervelsen. Dette eliminerer celleklumper, der kan tilstoppe instrumentet. Når du er klar, skal du klikke på Sorter og OK for at starte sorteringen. Juster flowhastigheden for at holde hastigheden under 10.000 hændelser pr. sekund.BEMÆRK: Det forventede forhold mellem LKS+ og CD34+ myeloide forfædre er 1:3 for en voksen (8-12 uger gammel) C57BL/6J hunmus ved steady state. De målrettede sorterede cellenumre nås normalt inden for 30 minutter efter sortering. Kvalitetskontrol og optælling af sorterede cellerBEMÆRK: Dette trin skal kun udføres under pilotforsøget til optimering af prøveforberedelsestrin med det formål at teste renheden af de sorterede celler, der skal bruges til kerneisolering. Når protokollen er fuldt optimeret, anbefales det ikke at udføre dette kvalitetskontroltrin i opfølgningsforsøgene for at undgå unødvendigt spild af udgangsmateriale, der kan være tilgængeligt i lavt antal til kerneisolering.Renhedskontrol ved flowcytometriOverfør 10 μL af de sorterede celler til et nyt FACS-rør indeholdende 90 μL FACS-buffer. Hent og registrer data efter sortering for at kontrollere sorteringens renhed og levedygtighed. Sørg for, at mindst 95 % af cellerne vises i interesseporten, som defineret i 3-6 og illustreret i figur 3B. Kerneisolering fra sorterede knoglemarvs-HSPC’erBrug protokollen “Low Cell Input Nuclei Isolation” i tillægget fra 10X Genomics Demonstrated Protocol – Nuclei Isolation for Single Cell Multiome ATAC + GEX sequencing (CG000365 – Rev C)27, med følgende ændringer foretaget for at optimere kernegendannelse:Lysistid: Kør et piloteksperiment for denne protokol for at identificere den bedste lysistid for kerneisolering. Sørg for at opnå en komplet cellelyse, samtidig med at intakte kerner opretholdes.BEMÆRK: Trin f i ovennævnte 10X Genomics-protokol27 instruerer om at “inkubere [i lysisbuffer] i 3-5 minutter på is”. Under pilotforsøget testes mindst 3 min, 4 minutter og 5 minutter og vurderes den genvundne kernemængde ved at tælle og kvalitet ved flowcytometri og mikroskopibilleddannelse for at vælge den optimale lysisvarighed (se beskrivelsen af disse kvalitetskontrolkontroller nedenfor). For at skåne reagenser skal du erstatte den fortyndede kernebuffer med PBS 0,04% BSA i pilotforsøget. For knoglemarv HSPC’er blev 3 min identificeret som den optimale lysisvarighed. Cellecentrifugeringer: Ved alle cellesuspensionscentrifugeringer centrifugeres ved 300 x g i 7 minutter (i stedet for 5 minutter ved CG000365 – Rev C)27 ved 4 °C. Kernecentrifugeringer: Udfør alle centrifugeringer af kerneopslæmning ved 500 x g i 5 minutter pr. CG000365 – Rev C27. Kerneopsamling: I trin b, efter resuspendering i 50 μL PBS 0,04% BSA og overførsel til et 0,2 ml rør, tilsættes 50 μL PBS 0,04% BSA til det originale rør og pipetteblanding for at opsamle eventuelle resterende celler. Overfør til 0,2 ml røret for at nå et samlet volumen på 100 μL. Fremover vil det samlede volumen være 100 μL i stedet for protokollens 50 μL. Nedstrømstrinnene justeres i overensstemmelse hermed (f.eks. fjernes 90 μL i stedet for 45 μL for trin d; for trin e tilsættes 90 μL lysisbuffer i stedet for 45 μL). I trin m resuspenderes kernepillen i 12 μL fortyndet kernebuffer i stedet for 7 μL. Tæl de isolerede kerner. I et tomt 0,5 ml rør tilsættes 10 μL 0,4% trypanblå og 8 μL PBS 0,04% BSA. Der tilsættes 2 μL kerner til røret, og tæl kernerne som beskrevet i 1.3.8. Brug en automatiseret celletæller efter leverandørens anbefalinger. Renhedskontrol ved flowcytometriBEMÆRK: Dette trin skal kun udføres under pilotforsøget til optimering af prøveforberedelsestrin for at teste renheden af kernerne, der skal lægges på 10X Chromium-chippen. Når protokollen er fuldt optimeret, anbefales det ikke at udføre dette kvalitetskontroltrin i opfølgningseksperimenterne for at undgå unødvendigt spild af indsamlede kerner, der kan være tilgængelige i lavt antal.Efter fuldstændig kerneisolering overføres 6 μL kerneresuspension i et nyt FACS-rør, der er fyldt med 150 μL FACS-buffer. Tilsæt 3 μL 7-AAD og inkuber i 5 minutter på is. Hent og registrer data efter sortering for at kontrollere sorteringens renhed og levedygtighed. Sørg for, at mindst 95 % af kernerne vises i interesseporten som defineret i protokol 1, trin 4.2 (se figur 4). Kvalitetskontrol af oprensede kerner ved mikroskopi:BEMÆRK: Dette trin skal kun udføres under pilotforsøget til optimering af prøveforberedelsestrin for at teste kvaliteten af kernerne, der skal indlæses på 10X Chromium-chippen. Når protokollen er fuldt optimeret, anbefales det ikke at udføre dette kvalitetskontroltrin i opfølgningseksperimenterne for at undgå unødvendigt spild af indsamlede kerner, der kan være tilgængelige i lavt antal.Fortsæt som beskrevet i trin 1.5.3. Udfør multiome-analyseFortsæt straks til Chromium Next GEM Single Cell Multiome ATAC + Gene Expression User Guide (CG000338 – Rev F)25.