A Set of In Situ Informed Simulated Medium Formats for Culturing Environmentally Acquired Anaerobic Microorganisms
Summary
The focus of this paper is to detail best practices for making media for fastidious anaerobic microorganisms acquired from an environment. These methods help manage anaerobic cultures and can be applied to support the growth of elusive uncultured microorganisms, the “microbial dark matter.”
Abstract
Culture-dependent research of anaerobic microorganisms rests upon methodological competence. These methods must create and maintain suitable growth conditions (e.g., pH and carbon sources) for anaerobic microorganisms while also allowing samples to be extracted without compromising the artificial environment. To this end, methods that are informed by and simulate an in situ environment can be of great aid in culturing microorganisms from that environment. Here, we outline an in situ informed and simulated anaerobic method for culturing terrestrial surface and subsurface microorganisms, emphasizing anaerobic sample collection with minimal perturbation. This protocol details the production of a customizable anaerobic liquid medium, and the environmental acquisition and in vitro growth of anaerobic microorganisms. The protocol also covers critical components of an anaerobic bioreactor used for environmental simulations of sediment and anaerobic liquid media for environmentally acquired cultures. We have included preliminary Next Generation Sequencing data from a maintained microbiome over the lifespan of a bioreactor where the active culture dynamically adjusted in response to an experimental carbon source.
Introduction
Most microorganisms remain uncultured; this is supported by the great disparity between cells observed through microscopy contrasted by the few microorganisms successfully cultured using agar plates. Staley and Konopka named this disparity the "Great Plate Count Anomaly"1. The estimated unaccounted diversity is supported by metagenomic and metatranscriptomic data showing many novel genera distributed in rank abundance curves from several different environments2. Microorganisms that have been observed (generally by random shotgun sequencing of a microbial community) but have not been cultured have been referred to as "microbial dark matter"3,4.
In the age of -omics, culturing microorganisms remains imperative to fully evaluate genomic data and verify the function/phenotype of genes present. Sequencing cultured microorganisms is still the only way to confidently obtain complete genomes until technologies such as shotgun metagenomics and metagenome-assembled genomes from the environment become admissibly infallible5. Genomic evaluations coupled with cultured microorganisms provide strong inferences for understanding "microbial dark matter." Many members of the "microbial dark matter" perform crucial functions that impact the cycling of nutrients and other elements and the production of valuable natural products, support ecological systems, and perform ecological services. From the medical perspective, about half of all currently marketed pharmaceuticals are products and derivatives of products from bacteria, and profiling uncultured species is suspected to reveal the antibiotics of the future. To gain access to this uncultured majority, a variety of culturing methodologies must be increased6. Among the members of the "microbial dark matter," anaerobic oligotrophic microorganisms are largely underreported and likely hold ecologically and industrially valuable biochemical pathways7, making them important targets of culturing. However, anaerobic oligotrophic microorganisms are more difficult to culture than their aerobic and copiotrophic counterparts due to often-required longer incubation times, fastidious conditions (e.g., particular non-standard in vitro temperatures), and the use of specialized media recipes.
Current developing techniques to culture members of the "microbial dark matter," including novel anaerobic oligotrophic microorganisms, have greatly improved our understanding and increased the representation of these microorganisms within the phylogenetic tree. Current techniques using informed media for culturing novel microorganisms (i.e., media which is derived using knowledge of the microorganism/s of interest) can be separated into three distinct methods. The first of the methods entails the direct removal of a discrete section of the environment for transfer into an in vitro growth chamber that already contains the microorganisms of interest within a membrane. The discrete section (e.g., seawater) acts to provide the microorganisms of interest with the geochemical habitat they use in situ, while the membrane arrests the movement of cells across (cells of interest will remain within; extraneous cells that arrived with the discrete section will remain without). By including compounds naturally available to target microorganisms in their natural habitat, such microorganisms can be cultured8. The second method utilizes metatranscriptomics or genomics to elucidate metabolic capabilities, providing clues to narrow culturing parameters for a targeted medium design. This approach provides an eco-physiological profile that can be used to target the enrichment of specific types of microorganisms out of an environment. The medium's provisions are catered to the identified genes present that are presumed to support the targeted microorganism(s) to reduce enrichment diversity,9,10. One caveat is that genomic information does not directly infer the expression of genes, while transcriptomic information does.
The third method encompasses environmentally informed and simulated media, distinct from the first method, which does not simulate the media but rather uses the environment directly as a source of media. This third method requires environmental reconnaissance of the geochemistry of a field site containing microorganisms of interest. With this knowledge, primary components and physical parameters are identified to produce an environmentally informed simulated medium. The medium then receives a direct infusion of microorganism-containing sediment or liquid from the environment into the medium. This method is of particular value in cases where the culturing microbiologist does not have access to sufficient quantities of source environment (as needed for the first method) nor appropriate metatranscriptomic or genomic data (as needed for the second).
The following protocol is an example of the third method; it is informed by and aims to simulate environments of interest. Three naïve media recipes targeting different anaerobic microorganism cultures acquired in the field are presented in parallel within the protocol. The three cultures represented are mixed cultures originating from soil (hereafter, soil mixed culture), mixed cultures originating from within a borehole (hereafter, borehole mixed culture), and an isolated methanogen originating from within a borehole (hereafter, borehole isolated methanogen). The compound identities and amounts in the media recipes shared here are meant as a beginning guide; they are able and encouraged to be customized to the reader's environment and microorganisms of interest.
Protocol
1. Production of customizable anaerobic liquid medium
- Medium for culture bottles (production of 500 mL)
- Measure and add compounds to a 1 L bottle and adjust the pH using the column in Table 1 corresponding to the reader's culture of interest (amounts in Table 1 are recorded for production of 1,000 mL, adjust accordingly). Mix compounds to homogeneity by swirling the bottle.
- Heat the liquid to boiling by microwaving the bottle for 5-6 min. Open the microwave often and swirl the liquid gently using a heat-resistant glove. Open the microwave quickly if the liquid goes still after bubbling; otherwise, the liquid may rapidly expand and overflow the bottle.
- Attach a sterile 10 mL glass pipette to a N2 gas manifold setup (based on the design of Balch et al.11, though not requiring a heated copper column for N2 gas of purity; see Supplemental File 1 for more gas manifold information). Open the manifold to vent O2 and allow N2 gas to flow out.
- Place the pipette tip into the liquid and adjust the flow of gas so that bubbles are moderately vigorous. Flush the liquid with N2 until the bottle is no longer too hot to touch.
NOTE: This is dependent on the contents of the media but generally takes ~20 min. - While waiting for the bottle to cool, set out 10 sterile 100 mL culture bottles; or if preparing media for collection of soil mixed culture, set out two sterile 500 mL bottles.
- Once the media bottle is cool to the touch, pull the pipette tip up into the headspace of the bottle. Add 0.125 g of NaHCO3 and 0.300 g of Na2S∙9 H2O and wait until resazurin turns colorless.
CAUTION: Na2S∙9 H2O is corrosive, toxic, and an irritant. Use personal protective equipment when handling and rinse metal tools thoroughly after contact. - While waiting for resazurin to change color, attach metal cannulas to the gas manifold setup and place the cannula tips into culture bottles. Adjust the flow of N2 into the bottles to a moderate rate, so that the liquid added in the subsequent step does not slosh about due to gas flow.
- Once resazurin is colorless, aliquot 50 mL of liquid into each culture bottle or 250 mL of liquid into each 500 mL bottle.
- Cap each bottle with a matching-sized rubber stopper immediately after removing the cannulas. Secure the stopper with an aluminum seal (for culture bottles) or a GL45 open-top screw cap (for 500 mL bottles).
- Place the bottles in an autoclavable bin and fill the bin with tap water until the water levels match those in the bottles.
NOTE: This will ensure the bottles do not rupture due to stress caused by temperature differentials on the glass while autoclaving. - Autoclave the bottles at 121 °C for 30 min.
- Medium for bioreactor (production of 8 L)
- Preparation of carboy
- Open the valve of a 1/4" (6.4 mm) hose barb ball valve. Attach two 6 cm lengths of 1/4" (6.4 mm) inner diameter (ID) and 1/2" (12.7 mm) outer diameter (OD) silicone tubing onto either end of the hose barb ball valve.
- Attach to one end of the assembly a 5/16"-1/4" (7.9 mm-6.4 mm) hose barb adapter fitting. Attach the other end of the assembly to the hose barb of an 8 L carboy.
- Use a zip tie to secure the connection between the carboy's hose barb and the tubing, and the connection between the hose barb adapter fitting and the tubing.
- Autoclave the carboy with assembly at 121 °C for 30 min.
- Medium production
- Close the valve of the hose barb ball valve on the 8 L carboy assembly.
- Measure and add compounds to the 8 L carboy and adjust the pH using the column in Table 1 corresponding to the reader's culture of interest (amounts in Table 1 are recorded for production of 1,000 mL, adjust accordingly). Add a stir bar and use a stirring hot plate to mix the compounds to homogeneity.
- Heat the liquid to boiling with the stirring hot plate. Allow the liquid to boil for 30 min.
NOTE: Heating the liquid to boiling is dependent on the contents of the media but can take 2-3 h. - Attach a sterile 10 mL glass pipette to a N2 gas manifold setup (based on the design of Balch et al.11, though not requiring a heated copper column for N2 gas of purity; see Supplemental File 1 for more gas manifold information). Open the manifold to vent O2 and allow N2 gas to flow out.
- Turn off the heat, then place the pipette tip into the liquid, and adjust the flow of gas so that the bubbles are moderately vigorous but not overflowing. Flush the liquid with N2 for 30 min.
- Pull the pipette tip up into the headspace of the carboy. Add 2.0 g of NaHCO3 and 4.8 g of Na2S∙9 H2O, and wait until resazurin turns colorless.
CAUTION: Na2S∙9 H2O is corrosive, toxic, and an irritant. Use personal protective equipment when handling and rinse metal tools thoroughly after contact. - Once resazurin is colorless, remove the glass pipette and immediately cap the carboy with a #10 stopper. Twist a wire over the stopper and around the lip of the carboy to secure the stopper.
- Preparation of carboy
2. in situ acquisition of environmental anaerobic microorganisms
- Surface anaerobic sample acquisition (soil mixed culture)
- Bring the bottle/s of mixed soil culture media made as above and a corer into the field.
NOTE: A Giddings 2 5/8" standard taper soil bit is used by the authors (Table of Materials). However, any soil corer or trowel that can sample to the desired depth may be used. - Push (pile/stake drive) the corer into the sediment until the top of the sample collection cylinder is flush with the surface of the sediment.
- Twist the corer 90° to free the core and pull upward until the sample is extracted from the environment.
- In loose sediments (such as what may be encountered when sampling wetlands), cover the base of the core with a new nitrile gloved hand as it is being extracted to prevent the sample from falling out or disseminating into the water on extraction.
- Quickly approximate by eye 6-7 cm up from the base of the core. Use a new nitrile gloved hand to slice into the core and separate the bottom 6-7 cm.
- Immediately transfer the bottom of the core to the culture bottle. If the soil/sediment is too large, quickly form it to fit into the bottle easier, minimizing exposure to the aerobic atmosphere as much as possible.
- Once the sample is transferred, immediately reposition the stopper and hold it in place with a screw cap.
- Keep the sample cool. Refrigerate the collection bottle at 4 °C upon return to the lab.
- Bring the bottle/s of mixed soil culture media made as above and a corer into the field.
- Subsurface anaerobic sample acquisition (borehole mixed culture)
- Set out a sterile 1 L bottle by the gas manifold. Cap the bottle with a rubber stopper. Drip 100% ethanol onto the stopper and set it on fire using a lighter.
- Attach a sterile 23 G needle to the N2 gas manifold setup. Open the manifold to vent O2 and allow N2 (or Ar) gas to flow out at a moderate rate.
- Once the stopper is no longer on fire, insert the needle into the bottle. Insert a second sterile 23 G needle not attached to the gas manifold into the bottle. Flush the bottle for 1-2 min.
- Extract the needle not attached to the gas manifold. Pressurize the bottle for 1 min. Extract the remaining needle.
- Bring the pressurized bottle and a sterilized 23 G needle to the borehole site. Draw up liquid from the borehole following the methods of Merino et al.12.
NOTE: Depending on the method, connection, and flow rate of the borehole, this changes the strategy of acquiring a water sample. If there is a continuous flow of water, collecting water as outlined in step 2.2.6. will be sufficient. If the flow of water is batch collected or low flow, then an alternative inlet/outlet method can be used. In all situations, minimize contamination, and attempt to ensure that the sample collected is representative of the environment desired to sample. This is a broadly generalized statement because it has been the authors' experience that every sample collection event from government or private sites had to be highly adaptable to the available accessibility. - Quickly open the bottle, pump in liquid to fill the bottle fully, and close the bottle.
- Insert the 23 G needle that was brought along to the site into the bottle's rubber stopper to alleviate pressure in the bottle. Extract the needle.
- Keep the sample cool. Refrigerate the collection bottle at 4 °C upon return to the lab.
3. in vitro growth of anaerobic microorganisms acquired in the field
- Growth by culture bottle
- Set out a stoppered sterile bottle of N2 and the culture bottles and collection bottle prepared as above. Drip 100% ethanol onto the stoppers and set them on fire using a lighter.
- Assemble a 23 G needle onto a 1 mL syringe. Once the stoppers are no longer on fire, insert the needle into the N2 bottle and draw up ~1 mL of N2. Extract the needle.
- Slowly press down on the plunger to release the N2. As soon as the syringe is empty, insert the needle into the collection bottle and draw up 0.5 mL of the environmental sample. Extract the needle.
- Insert the needle into the culture bottle and inject the sample. Extract the needle.
- Swirl the bottle and place it in an incubator at temperature, as informed by Table 1 or the reader's environment of interest.
NOTE: Culture progress (either bottle or bioreactor) is monitored by using a 0.02 mm deep Petroff-Hauser hemocytometer. Enrichments of anaerobic or fastidious microorganisms typically do not reach cell densities where OD600 is usable, and other detection methods are impractical for routine and repetitive monitoring. Detection limits, quality control, and statistical calculations are dependent on the type of cell counting chamber and the user's needs.
- Growth by bioreactor
- Preparation of a balloon with a modified syringe
- Remove the plunger from three 10 mL luer lock syringes (one for each final assembly). Cut off the flange from each syringe and file the severed end so it will not pierce the balloons. Clean all filed chips out of the modified syringe.
- Prepare a doubled balloon by placing one balloon inside another.
- Stretch the end of the doubled balloon over the modified 10 mL syringe.
- Slightly separate the end of the outer balloon from the inner balloon. Squeeze out all the air trapped between the balloons.
- Tighten a zip tie around the base of the balloons to secure the balloons to the syringe.
- Preparation of a modified stopper
- Remove the plungers from ten 1 mL luer lock syringes (two for each stopper to be modified). Cut off the flange from the syringes.
- Set out two rubber stoppers size #10 (for 8 L carboys) and three sized for bottles with GL45 thread finish (for catch bottles). Using a 1/4" (6.4 mm) drill bit, drill two holes through the top of each rubber stopper wide enough for the modified syringes to fit and narrow enough for the fit to be airtight.
- Insert the modified syringes through the holes of the rubber stopper.
- Clean and autoclave the modified stopper at 121 °C for 30 min.
- Preparation of the bioreactor assembly
- Disinfect with 70% ethanol all stopcocks, female luer lock adapter connectors, and other non-autoclavable bioreactor materials, and autoclave at 121 °C for 30 min all tubing (after cutting to length), stoppers, glass wool, and other autoclavable bioreactor materials.
- Set a sterile bioreactor (Figure 1 and Figure 2) on a ring stand and secure it with a chain strap. Also set out a water bath, a peristaltic pump, the stoppered 8 L carboy containing medium (from protocol step 1.2), one 3.5 L bottle (non-sampling catch bottle), and one 500 mL bottle (sampling catch bottle).
- Stoppering the carboy
- Set out a balloon with a modified syringe. Connect a three-way stopcock to the syringe's luer lock tip.
- Take the balloon assembly and connect the three-way stopcock to the tubing of a tank of N2. Turn the three-way stopcock to block off the end open to the atmosphere.
- Open the gas tank and fill the balloon with N2. Once full, turn the three-way stopcock to block off the end to the balloon. Close the gas tank and disconnect the tank tubing from the three-way stopcock.
- Connect the following in sequence: a three-way stopcock (of the balloon with the modified syringe), a two-way stopcock, a female luer lock adapter coupler, and the luer lock tip of one of the modified stopper's syringes.
- To the other syringe's luer lock tip, connect a two-way stopcock. Close both valves of the two-way stopcocks on the modified stopper assembly.
- Untwist the wire holding the carboy's unmodified size #10 stopper. Quickly replace the unmodified stopper with the modified stopper assembly and twist the wire as before to secure the modified stopper assembly to the carboy.
NOTE: If confident and prepared, the modified stopper assembly can be used to stopper the carboy at the final step of medium preparation (step 1.2.2.7). - Turn the three-way stopcock to block off the end open to the atmosphere. Open the in-sequence two-way stopcock. The gas of the balloon and the headspace of the 8 L carboy are now connected.
- Stoppering the non-sampling and sampling catch bottles
- Stopper the 3.5 L non-sampling catch bottle with a modified stopper sized for bottles with GL45 thread finish, applying 70% ethanol to the base of the stopper to help fit it into the bottle. Cap the bottle with a GL45 open-top screw cap.
- Connect the following in sequence: a balloon with the modified syringe's luer lock tip, a three-way stopcock, a female luer lock adapter coupler, and the luer lock tip of one of the modified stopper's syringes. Turn the three-way stopcock to block off the end open directly to the atmosphere.
- To the other syringe's luer lock tip, connect a female luer lock adapter coupler.
- Repeat the above three steps for the 500 mL sampling catch bottle.
- Setup of tubing
- Measure the distance between the bioreactor water jacket ports and the water bath hose barbs. Cut two lengths of 3/16" (4.8 mm) ID, 3/8" (9.5 mm) OD silicone tubing to span this distance.
- Assemble two straight hose connectors with GL14 open-top screw caps. Attach each to one end of the two tubing pieces.
- Twist the screw caps onto the ports of the bioreactor water jacket. Attach the other ends of the tubing to the water bath hose barbs so that the water will flow out of the water bath, enter at the bottom of the bioreactor water jacket, exit at the top of the water jacket, and return to the water bath. Tighten zip ties around the ends of the tubing to secure the tubing to the water bath hose barbs.
- For pumps similar to the VWR Ultra Low Flow Variable Flow Mini-Pump, thread the accompanying 3/16" (4.8 mm) OD slotted tubing assembly through the pump.
- Measure the distance between the carboy tubing assembly and the peristaltic pump tubing assembly. Cut a length of 3/16" (4.8 mm) ID, 3/8" (9.5 mm) OD silicone tubing to span this distance.
- Attach one end of the tubing to the hose barb of the carboy tubing assembly and the other to the hose barb of the peristaltic pump tubing assembly, oriented so that the medium will flow from the carboy and through the peristaltic pump.
- Measure the distance between the peristaltic pump tubing assembly and the bottom port of the bioreactor. Cut a length of 3/16" (4.8 mm) ID, 3/8" (9.5 mm) OD silicone tubing to span this distance.
- Attach one end of this tubing to the hose barb of the peristaltic pump tubing assembly and carefully attach the other to the bottom port of the bioreactor. Taking care not to crack the glass but also to secure the connection, tighten a zip tie around the end of the tubing to secure the tubing to the bottom port of the bioreactor.
- Measure and cut approximately 5" (127mm) length of 3/16" (4.8 mm) ID, 3/8" (9.5 mm) OD silicone tubing.
- Attach a female luer lock adapter connector to a three-way stopcock. Attach any end of the three-way stopcock to the tubing.
- Assemble one angled hose connector with GL14 open-top screw cap. Attach it to the other end of the tubing.
- Twist the screw cap onto the topmost vertical port of the bioreactor. Turn the three-way stopcock to block off the end pointing to the bioreactor.
- Measure the distance between the topmost vertical port assembly and the non-sampling catch bottle assembly. Cut a length of 3/16" (4.8 mm) ID, 3/8" (9.5 mm) OD silicone tubing to span this distance. Cut the flanges off two 1 mL luer lock syringes and insert them into the ends of the tubing.
- Attach one end of this tubing assembly to the three-way stopcock of the topmost vertical port assembly and attach the other end to the female luer lock adapter connector on the non-sampling catch bottle assembly.
- Measure the distance between the topmost vertical port assembly and the sampling catch bottle assembly. Cut a length of 3/16" (4.8 mm) ID, 3/8" (9.5 mm) OD silicone tubing to span this distance. Cut the flanges off two 1 mL luer lock syringes and insert them into the ends of the tubing.
- Attach one end of this tubing assembly to the three-way stopcock of the topmost vertical port assembly and attach the other to the female luer lock adapter connector on the sampling catch bottle assembly.
- Filling the bioreactor
- Cap one of the bioreactor's GL18 ports with a white rubber septum. Cap the other GL18 port and remaining vertical exposed GL14 ports with GL18/GL14 sized PTFE faced silicone septa (PTFE facing the glass) and open top screw caps.
- With a 50 cm length rod, pack 0.9 g of glass wool into the base of the bioreactor.
- Autoclave 1.3 L of sand at 121 °C for 30 min. Fill the bioreactor with the sand using a funnel.
NOTE: Other sediment types informed by the reader's environment and microorganisms of interest can be substituted here. - Place the tubing to a tank of N2 into the top of the bioreactor and flush the bioreactor headspace with N2 for 2 min. Immediately cap the bioreactor with a #7-sized stopper and a GL45 open-top screw cap.
- From the non-sampling catch bottle, disconnect the tubing and turn the three-way stopcock to block off the end pointing to the balloon. Connect the tubing to a tank of N2 to the female luer lock adapter connector and flush the bioreactor headspace with N2 for 2 min. Immediately reconnect the tubing and return the three-way stopcock to block off the end open to the atmosphere.
- Repeat the above step for the sampling catch bottle.
- Turn the three-way stopcock of the bioreactor's topmost vertical port to block off the end pointing to the sampling catch bottle. Pump liquid from the carboy into the bioreactor. Pause the pump, and vent the sampling bottle's balloon, and fill the carboy's balloon with N2 as needed.
- Once the bioreactor has been filled with the medium, set the pump flow rate to 0.6 mL/min (digital value of 40 on the Mini-Pump).
NOTE: Other reader-desired flow rates can be substituted here. - Set the water bath temperature, as informed by Table 1 or the reader's environment of interest.
NOTE: Inoculum can be added at this point or later, depending on study specifics. If inoculating at this point, proceed to step 3.2.4.10. If inoculating later, proceed to step 3.2.5. - Set out a collection bottle or inoculated culture bottle and a stoppered sterile bottle of N2. Drip 100% ethanol onto the stoppers and set them on fire using a lighter.
- Assemble a 23 G needle onto a 60 mL syringe. Once the stoppers are no longer on fire, insert the needle into the N2 bottle and draw up ~50 mL of N2. Extract the needle.
- Slowly press down on the plunger to release the N2. As soon as the syringe is empty, insert the needle into the collection bottle and draw up 50 mL of the environmental sample. Extract the needle.
- Exchange the 23 G needle for a sterile long metal needle (cannula, length 31.5 cm). Insert the long metal needle through the white rubber septum on the bioreactor's GL18 port.
- Push the needle into the sediment and inject the inoculum. Extract the needle.
- Running bioreactor maintenance
- Each day that the bioreactor is running, perform the following:
- Check the water bath level. If low, add more water (use distilled water for equipment longevity by reducing corrosion and mineral buildup).
- Check the balloon of the non-sampling catch bottle; a full balloon will inhibit the flow of the medium. When full, turn the three-way stopcock to block off the end pointing to the catch bottle. Squeeze the balloon to empty it; then, return the three-way stopcock to block off the end open to the atmosphere; repeat until the balloon is empty.
- Check the balloon of the 8 L carboy. If low, close the in-sequence two-way stopcock and disconnect the three-way stopcock from the two-way stopcock. Refill and reattach the balloon assembly following steps 3.2.3.3.2, 3.2.3.3.3, and 3.2.3.3.7.
- Check the level of the medium in the 8 L carboy. If low, assemble another 8 L carboy and prepare more medium following protocol steps 1.2. and 3.2.3.3. Pause the pump, close the hose barb ball valve, quickly disconnect the old carboy, and connect the new carboy.
NOTE: For a flow rate of 0.6 mL/min, new medium will be needed every 9 days. - Check the level of the medium in the non-sampling catch bottle. If full, assemble another non-sampling catch bottle according to step 3.2.3.4.1. Flush the new bottle with N2, quickly disconnect the balloon with the modified syringe and tubing from the old bottle, and connect to the new bottle.
- Remove the modified stopper from a full non-sampling catch bottle. Autoclave the liquid at 121 °C for 30 min; then, dispose of the liquid according to the reader's institutional policies. Clean and autoclave all pieces of the non-sampling catch bottle assembly to prepare them for next use.
- Perform sampling every 7 (or reader-desired number of) days by doing the following:
- Turn the three-way stopcock of the bioreactor's topmost vertical port to block off the end pointing to the non-sampling catch bottle. Collect 250 mL of liquid.
NOTE: For a flow rate of 0.6 mL/min, collecting 250 mL will take ~7 h. - Return the three-way stopcock of the bioreactor's topmost vertical port to block off the end pointing to the sampling catch bottle. Disconnect the tubing and sampling catch bottle from the three-way stopcock of the bioreactor's topmost vertical port.
- After testing, storage, and/or proper disposal of sampling liquid, clean all pieces of the sampling catch bottle assembly except the balloon with the modified syringe. Autoclave all pieces at 121 °C for 30 min except the balloon with the modified syringe and female luer lock adapter connectors. Reassemble for the next sampling day.
NOTE: It is highly recommended to have duplicate or triplicate redundancy of parts, caps, tubing, and septa for rapid repair and replacement if necessary.
- Turn the three-way stopcock of the bioreactor's topmost vertical port to block off the end pointing to the non-sampling catch bottle. Collect 250 mL of liquid.
- Each day that the bioreactor is running, perform the following:
- Preparation of a balloon with a modified syringe
Representative Results
Here we show results from a bioreactor study using a borehole mixed culture medium preparation method and a bioreactor setup method as described herein. The borehole mixed culture medium was modified to contain as a carbon source a slurry of corn cobs processed by Oxidative Hydrothermal Dissolution (OHD)13,14. Modified borehole mixed culture medium was pumped into the bioreactor for 44 days at a rate of 0.4 mL/min. On day 23, an inoculum sourced from borehole BLM-1 in the Death Valley region of Nevada, USA, was added15. Liquid samples from the bioreactor were collected in a sampling catch bottle on days 1, 8, 15, 22, 23 (after inoculation), 30, 37, and 44.
To track the general status of the inoculum culture (and pre-inoculum autoclave-surviving denizens of the bioreactor sediment), bioreactor liquid samples were observed by direct cell counts. Direct cell counts of sampled liquid were done with a Petroff-Hauser hemocytometer. To learn the identity of microbial members of the bioreactor, DNA was extracted from bioreactor liquid samples, and the 16S rRNA genes were analyzed by Next Generation Sequencing (NGS). The NGS data were processed in-house using mothur following the MiSeq SOP16.
Before inoculation, direct cell counts were often below the detection limit (b.d.l.) of 1.0 × 106 cells/mL. Day 23 (after inoculation) had a low cell count of 2.6 × 106 cells/mL, while the following days 30, 37, and 44 had cell counts over 1.0 × 107 cells/mL (Figure 3). Although cell counts were b.d.l. on days 1, 8, and 22, DNA was extracted for NGS from every time point indicating a basal presence of pre-inoculum microorganisms in the sediment even after autoclaving. The major genus present (determined by the number of reads in each Operational Taxonomic Unit (OTU)) was stable on days 8, 15, and 22, and was identified as Geobacillus. After inoculation with the BLM-1 inoculum on day 23, Geobacillus no longer dominated, and new major genera arose along with a multitude of minorly present genera, which for the most part, maintained a presence to the end of the study (Figure 4). From these, we conclude the BLM-1 inoculum was an active and dynamically shifting culture within the bioreactor.
An additional study of the NGS data revealed members of the "microbial dark matter." OTUs with assigned taxonomic names containing the words "unclassified" or "uncultured" were used as a proxy for OTUs containing members of the "microbial dark matter." NGS data were gathered for day 44 (the final day of post-inoculation with microorganisms from borehole BLM-1) and filtered to not contain OTUs that had reads in pre-inoculation data (days 1, 8, 15, or 22) as these OTUs were not likely to have been in the inoculum. The filtered day 44 NGS data contained 1,844 OTUs and 3,396 reads. Of these, 366 OTUs (20%) and 925 reads (27%) were unclassified at the phylum level, and 1252 OTUs (68%) and 2357 reads (69%) were unclassified or uncultured at the genus level. Though short-term, by this proxy, this pilot study supports the potential of a bioreactor with environmentally informed media to culture members of the "microbial dark matter."
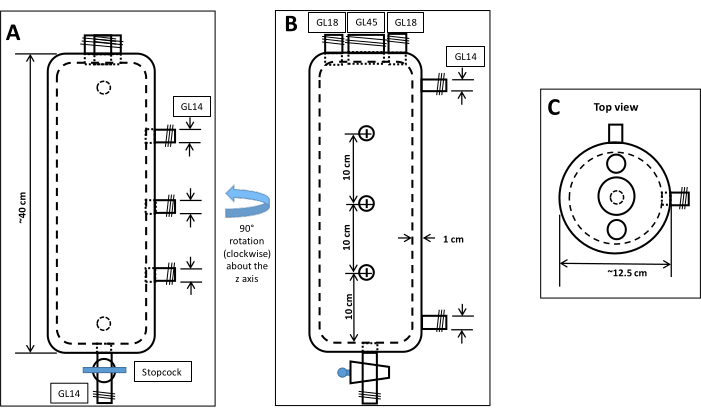
Figure 1: Schematic of custom-designed borosilicate bioreactor used for anaerobic culturing. (A) Bioreactor view from one side. (B) Rotating A by 90° (clockwise) about the z-axis gives the view). (C) A top view of the bioreactor. The jacketed design allows for temperature control with water or oil. The two jacket ports can be seen on the right side of the view in B. The inner chamber can be filled with sediment of any type or can be left empty of sediment for planktonic culturing. Three sampling ports (seen on the right side of the view in A) make it possible to remove material at different depths of the bioreactor column. Standard 45, 18, and 14 GL openings can be sealed using Teflon or butyl septa that will hold a gastight seal for 1-6 months. Please click here to view a larger version of this figure.

Figure 2: Active bioreactor assembly. Glassware presented in photo from left to right are (A) 8 L carboy, (B) bioreactor (see Figure 1), and (C) non-sampling catch bottle. Please click here to view a larger version of this figure.
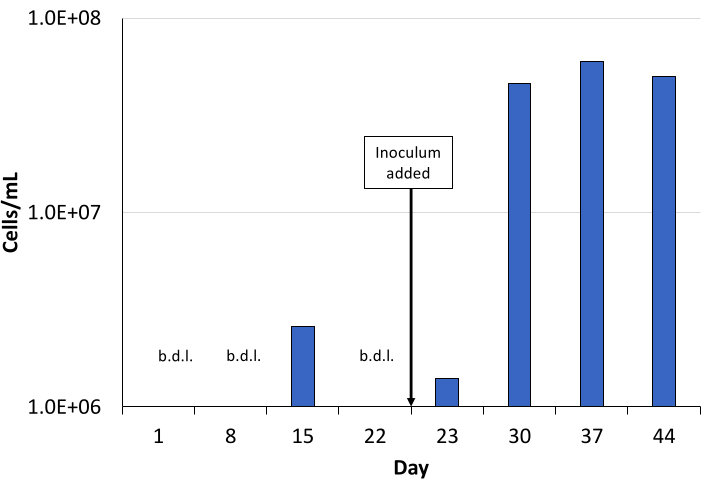
Figure 3: Direct cell counts from bioreactor study. Microbial populations within the bioreactor were monitored using a 0.02 mm Petroff-Hauser hemocytometer cell counting chamber. Inoculum was added at the start of day 23. Cell counts for days 1, 8, and 22 were below the detection limit. The effective detection limit of direct cell counting is 1 × 106 cells/mL. Abbreviation: b.d.l. = below the detection limit. Please click here to view a larger version of this figure.
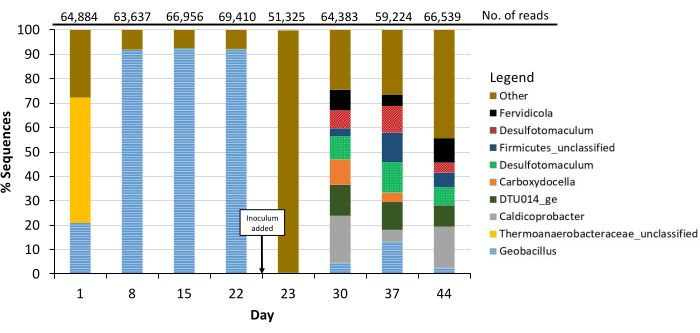
Figure 4: Bioreactor Next Generation Sequencing. OTUs are represented in stacked bar graphs indicating the percentage of reads belonging to unique OTUs from each time point of the bioreactor study. Unique OTUs were assigned to their associated taxonomy at a genus cutoff. Genus 'Other' is defined as all genera less than 10% of reads in each time point. The total number of DNA reads represented is 506,358. The number of reads for each time point is listed at the top of the graph. Inoculum was added at the start of day 23. Abbreviation: OTU = Operational Taxonomic Unit. Please click here to view a larger version of this figure.
| Compound | Amount for soil mixed culture | Amount for borehole mixed culture | Amount for borehole isolated methanogen |
| Distilled water | 1000.0 mL | 1000.0 mL | 1000.0 mL |
| HEPES | 3.600 g | – | – |
| MOPS | – | – | 2.000 g |
| MgCl2 ∙6 H2O | 0.400 g | 0.400 g | 0.400 g |
| KCl | 0.500 g | 0.250 g | 0.500 g |
| NH4Cl | 0.268 g | 0.268 g | 0.268 g |
| Na2SO4 | – | 0.284 g | 1.500 g |
| Na2HPO4 ∙2 H2O | – | – | 0.356 g |
| 1 M KH2PO4 at pH 6 | 1.0 mL | 1.0 mL | – |
| 1 M H3BO3 at pH 9.4 | 1.0 mL | 1.0 mL | 1.0 mL |
| Trace minerals | 1.0 mL | – | 1.0 mL |
| Vitamins | – | – | 1.0 mL |
| 1 mg/mL sodium resazurin | 0.4 mL | 0.3 mL | 0.4 mL |
| pH adjustment to | – | 7.0 | 7.0 |
| pH adjustment by | – | NaOH | KOH |
| Incubation temperature | 25 °C | 60 °C | 47 °C |
Table 1: Media composition. List of compounds and physical parameters for each medium. The presented media are naïve; they do not contain carbon and energy sources as these can be specific to and should be informed by the reader's microorganisms and environment of interest. See the discussion section for ideas of possible carbon and energy source additions.
Supplemental File 1: Gas manifold setup. Please click here to download this File.
Discussion
The medium production section of this protocol (section 1) owes its structure to the modified Hungate technique of Miller and Wolin17, which has been widely used since its publication. The practicality of this expanded protocol comes from its descriptive nature and pairing with the in situ acquisition of microorganisms. Culture bottles containing environmentally informed and simulated media have been used to successfully culture the following in situ-acquired former members of the "microbial dark matter": anaerobic subsurface bacteria Thermoanaerosceptrum fracticalcis strain DRI1318, Caldiatribacterium inferamans strain SIUC1 (accession number MT023787; manuscript in preparation), and Anaerothermus hephaesti strain SIUC3 (accession number MK618647; manuscript in preparation), and anaerobic subsurface bacterium uncharacterized strain DRI14 (accession number KR708540).
One focus of this protocol is that it is made to be highly adaptable to surface and subsurface anaerobic environments and microorganisms. First, the medium composition is encouraged to be altered to reflect the composition of the environment of interest. For example, 3,000 mg/L of NaCl can be added to the recipe when simulating an environment with ionic concentrations of 3,000 mg/L for both Na+ and Cl–; or 650 mL of sand and 650 mL of shale can be placed in the bioreactor when simulating an environment with a 1:1 volume ratio of sandstone to shale. Second, the medium composition can be modified to encourage the growth of specific microorganisms of interest. Particularly, naïve media like the three media in this paper are primed for and dependent upon such modifications. Examples of carbon and energy sources include fumarate18, peptone18, cellulose-containing organic matter19, acetate20, yeast extract19,20, xylitol (strain SIUC1), formate (strain SIUC3), and other potential sources such as H2/CO2, citrate, and glucose. Finally, the medium composition can be changed depending on the desired study. For example, the main interest of the study described in the representative results section was to discover how representative subsurface anaerobic microorganisms would respond to a unique mixed organic slurry (a slurry of corn cobs processed by OHD13,14); thus, its recipe contained this carbon source instead of more conventional options. Other potential amendments for specific studies include the addition of plastics, acids, and heavy metals, or other xenobiotics for bioremediation studies; the addition of recalcitrant carbon polymers such as cellulose and plastics for value-added depolymerized product studies; and the addition of manure and plant wastes for small-scale compost and other agricultural studies.
If post-reductant analysis of a medium is desired, such as measurements of pH, Oxidation Reduction Potential (ORP), and Dissolved Oxygen (DO), then this may be accomplished indirectly by sacrificing a representative number of medium bottles to confidently understand the character of that batch. Upon exposing a sealed medium bottle to air, it is strongly recommended that ORP and DO readings are taken under 1 min upon opening. Non-oxygen-dependent measurements (e.g., pH) can be taken non-urgently after the initial opening. Fastidious anaerobic microorganisms typically require ORP values < -200 mV for growth. If the medium results in an ORP value of > -200 mV and/or DO values >0.40 mg/L, additional Na2S (0.1-0.2 g/L) or another reductant (e.g., titanium citrate, L-cysteine, ascorbic acid) can be used to reduce the medium further.
Resazurin is often used as an oxygen indicator (as in this protocol); however, it is more accurately a redox indicator21, indicating the presence of oxygen indirectly as the oxygen changes the redox potential of a solution. Resazurin in solution initially appears blue in color. Upon exposure to reducing agents, the solution shifts irreversibly to a pink color. When the solution is further reduced, it reversibly shifts to colorless and returns to pink upon oxidization. The authors have observed instances where reduced media containing resazurin shifted from blue to pink but failed to shift to colorless. However, the authors have also observed that such media often shift to colorless after autoclaving, and that increased time spent boiling and flushing the media generally allows resazurin to shift to its colorless form after adding reductants. If desired, the concentration of oxygen in the medium can be verified with a dissolved oxygen probe.
All materials needed to run these bioreactors and the custom-made glass bioreactors themselves did not exceed ~$5K (this assumes support equipment is already available such as water baths, pumps, and incubators). These anaerobic cultivation techniques are economically friendly, as compared to commercial automated bioreactors which can start at >$10K. Additionally, many commercial bioreactors are switching to one-time-use reactor containers, which changes the dynamic of comparison and is not cost-effective for smaller research laboratories/universities to continually replace. Additionally, commercial systems may have manufacturing chemical biases (from the plastics used, grease on gaskets/fittings, or chemically treated water) that will affect the success of growing fastidious microorganisms. The low cost, customizability, and versatility to collect an anaerobic environmental sample, in combination with an informed media that can be inoculated, which results in generating enrichments for study, greatly enhances the chances of acquiring unique microorganisms.
Divulgazioni
The authors have nothing to disclose.
Acknowledgements
The authors would like to acknowledge the lineage of information and mentorship that has influenced/evolved these techniques over the years. Dr. Hamilton-Brehm as a former graduate student, postdoc, and current professor owes a debt of gratitude to those who took the time to teach anaerobic techniques: Dr. Mike Adams, Dr. Gerti Schut, Dr. Jim Elkins, Dr. Mircea Podar, Dr. Duane Moser, and Dr. Brian Hedlund. The Nature Conservancy and American Rivers supported this work through grants G21-026-CON-P and AR-CE21GOS373, respectively. Any opinions, findings, conclusions, or recommendations expressed in this paper are those of the authors and do not necessarily reflect the views of the Nature Conservancy or American Rivers. This work was supported by a grant from the SIU Advanced Energy Institute, which gratefully acknowledges funding awarded through the Advanced Energy Resource Board. NGS was performed by LC Sciences.
Materials
| General Materials | |||
| 1 L borosillicate bottle | Fisher Scientific | ||
| 1 mL syringe with slip tip | Fisher Scientific | ||
| 10 mL glass pipette | Fisher Scientific | ||
| 100 mL culture bottle | Fisher Scientifc | ||
| 20 mm hand crimper | Fisher Scientifc | ||
| 23 G needle | Fisher Scientifc | ||
| 500 mL borosilicate bottle | Fisher Scientific | ||
| Aluminum seal | Fisher Scientifc | ||
| Cannula, 31.5 cm length | Fisher Scientific | ||
| Cannula, 6 cm length | Fisher Scientifc | ||
| Corer | Giddings Machine Company | Assembled from company parts | |
| Gas manifold | Swagelok | Assembled from many different parts | |
| Lighter | Lowe's | ||
| N2 gas | Airgas | ||
| Nitrile gloves | Fisher Scientific | ||
| Rubber stopper (for GL45 bottles) | Glasgeratebau OCHS | ||
| Rubber stopper (for culture bottles) | Ace Glass | ||
| Stirring hot plate | Corning | ||
| Trace minerals | ATCC | ||
| Vitamins | ATCC | ||
| Bioreactor-specific Materials | |||
| #10 rubber stopper | Ace Glass | ||
| #7 rubber stopper | Fisher Scientifc | ||
| 1 mL syringe with luer lock tip | Fisher Scientifc | ||
| 1/4" hose barb ball valve | Amazon | ||
| 10 mL syringe with luer lock tip | Fisher Scientifc | ||
| 3.5 L borosilicate bottle | Fisher Scientific | ||
| 5/16" – 1/4" hose barb adapter fitting | Amazon | ||
| 60 mL syringe with luer lock tip | Fisher Scientifc | ||
| 8 L borosillicate carboy | Allen Glass | ||
| Angled hose connector for GL14 open top cap | Ace Glass | 7623-20 | |
| Balloon | Party City | ||
| Borosillicate bioreactor | Allen Scientific Glass | Custom made upon request | |
| Drill | Lowe's | ||
| Female luer lock adapter coupler | Amazon | ||
| GL14 open top cap | Ace Glass | 7621-04 | |
| GL18 open top cap | Ace Glass | 7621-08 | |
| GL45 open top cap | Ace Glass | ||
| PTFE faced silicone septum for GL14 open top cap | Ace Glass | 7625-06 | |
| PTFE faced silicone septum for GL18 open top cap | Ace Glass | 7625-07 | |
| Ring stand | Fisher Scientific | ||
| Ring stand chain clamp | Amazon | ||
| Ring stand clamp | Fisher Scientific | ||
| Silicone tubing; 1/4" id, 1/2" od | Grainger | 55YG13 | |
| Silicone tubing; 3/16" id, 3/8" od | Grainger | ||
| Straight hose connector for GL14 open top cap | Ace Glass | 7623-22 | |
| Three-way stopcock | Amazon | ||
| Two-way stopcock | Amazon | ||
| Ultra low flow variable flow mini-pump | VWR | ||
| Water bath | Fisher Scientifc | ||
| White rubber septum for 13-18 mm od tubes | Ace Glass | 9096-49 | |
| Wire | Lowe's | ||
| Zip tie | Lowe's |
Riferimenti
- Staley, J. T., Konopka, A. Measurement of in situ activities of nonphotosynthetic microorganisms in aquatic and terrestrial habitats. Annu. Rev. Microbiol. 39 (1), 321-346 (1985).
- Lloyd, K. G., Steen, A. D., Ladau, J., Yin, J., Crosby, L. Phylogenetically novel uncultured microbial cells dominate earth microbiomes. MSystems. 3 (5), e00055 (2018).
- Rinke, C., et al. Insights into the phylogeny and coding potential of microbial dark matter. Nature. 499 (7459), 431-437 (2013).
- Marcy, Y., et al. Dissecting biological "dark matter" with single-cell genetic analysis of rare and uncultivated tm7 microbes from the human mouth. Proc. Natl. Acad. Sci. USA. 104 (29), 11889-11894 (2007).
- Giovannoni, S., Stingl, U. The importance of culturing bacterioplankton in the’omics’ age. Nat. Rev. Microbiol. 5 (10), 820-826 (2007).
- Stewart, E. J. Growing unculturable bacteria. J. Bacteriol. 194 (16), 4151-4160 (2012).
- Dedysh, S. N. Cultivating uncultured bacteria from northern wetlands: Knowledge gained and remaining gaps. Front. Microbiol. 2, 184 (2011).
- Kaeberlein, T., Lewis, K., Epstein, S. S. Isolating" uncultivable" microorganisms in pure culture in a simulated natural environment. Science. 296 (5570), 1127-1129 (2002).
- Bomar, L., Maltz, M., Colston, S., Graf, J. Directed culturing of microorganisms using metatranscriptomics. mBio. 2 (2), e00012 (2011).
- Tyson, G. W., et al. Genome-directed isolation of the key nitrogen fixer Leptospirillum ferrodiazotrophum sp. nov. from an acidophilic microbial community. Appl. Environ. Microbiol. 71 (10), 6319-6324 (2005).
- Balch, W. E., Fox, G. E., Magrum, L. J., Woese, C. R., Wolfe, R. S. Methanogens: Reevaluation of a unique biological group. Microbiol. Rev. 43 (2), 260-296 (1979).
- Merino, N., et al. Subsurface microbial communities as a tool for characterizing regional-scale groundwater flow. Sci. Total Environ. 842, 156768 (2022).
- . Process for the dissolution of coal, biomass and other organic solids in superheated water. U.S. patent 8,563,791 Available from: https://patentimages.storage.googleapis.com/23/49/97/e50f70357c62d8/US8563791.pdf (2013)
- . Production of organic materials using oxidative hydrothermal dissolution. U.S. patent 10,023,512 Available from: https://patentimages.storage.googleapis.com/a6/28/30/7f14a156b44bdf/US10023512.pdf (2018)
- Mullin, S. W., et al. Patterns of in situ mineral colonization by microorganisms in a~ 60 c deep continental subsurface aquifer. Front. Microbiol. 11, 536535 (2020).
- Kozich, J. J., Westcott, S. L., Baxter, N. T., Highlander, S. K., Schloss, P. D. Development of a dual-index sequencing strategy and curation pipeline for analyzing amplicon sequence data on the miseq illumina sequencing platform. Appl. Environ. Microbiol. 79 (17), 5112-5120 (2013).
- Miller, T. L., Wolin, M. A serum bottle modification of the hungate technique for cultivating obligate anaerobes. Appl Microbiol. 27 (5), 985-987 (1974).
- Hamilton-Brehm, S. D., et al. Thermoanaerosceptrum fracticalcis gen. nov. sp. nov., a novel fumarate-fermenting microorganism from a deep fractured carbonate aquifer of the us great basin. Front. Microbiol. 10, 2224 (2019).
- Hamilton-Brehm, S. D., et al. Caldicellulosiruptor obsidiansis sp. nov., an anaerobic, extremely thermophilic, cellulolytic bacterium isolated from obsidian pool, yellowstone national park. Appl. Environ. Microbiol. 76 (4), 1014-1020 (2010).
- Hamilton-Brehm, S. D., et al. Thermodesulfobacterium geofontis sp. nov., a hyperthermophilic, sulfate-reducing bacterium isolated from obsidian pool, yellowstone national park. Extremophiles. 17 (2), 251-263 (2013).
- Twigg, R. S. Oxidation-reduction aspects of resazurin. Nature. 155 (3935), 401-402 (1945).
