Området for bottom-up syntetisk biologi fokuserer på konstruktion af (minimale) celler 1,2 og metaboliske bioreaktorer til bioteknologiske 3,4 eller biomedicinske formål 5,6,7,8. Konstruktionen af syntetiske celler giver en unik platform, der giver forskere mulighed for at studere (membran) proteiner under veldefinerede forhold, der efterligner dem i native miljøer, hvilket muliggør opdagelsen af emergente egenskaber og skjulte biokemiske funktioner af proteiner og reaktionsnetværk9. Som et mellemliggende skridt mod en autonomt fungerende syntetisk celle udvikles moduler, der fanger væsentlige træk ved levende celler såsom metabolisk energibevarelse, protein- og lipidsyntese og homeostase. Sådanne moduler forbedrer ikke kun vores forståelse af livet, men har også potentielle anvendelser inden for medicin8 og bioteknologi10.
Transmembranproteiner er kernen i stort set ethvert metabolisk netværk, da de transporterer molekyler ind eller ud af cellen, signalerer og reagerer på miljøets kvalitet og spiller adskillige biosyntetiske roller. Således kræver konstruktionen af metaboliske moduler i syntetiske celler i de fleste tilfælde rekonstituering af integrerede og/eller perifere membranproteiner til et membrandobbeltlag sammensat af specifikke lipider og høj integritet (lav permeabilitet). Håndteringen af disse membranproteiner er udfordrende og kræver specifik viden og eksperimentelle færdigheder.
Der er udviklet flere metoder til at rekonstituere membranproteiner i fosfolipidvesikler, oftest med det formål at studere funktionen11,12, regulering13, kinetiske egenskaber14,15, lipidafhængighed15,16 og/eller stabilitet17 af et specifikt protein. Disse metoder omfatter hurtig fortynding af vaskemiddelopløseligt protein til vandige medier i nærvær af lipider18, fjernelse af vaske- og rengøringsmidler ved inkubation af vaskemiddelopløseligt protein med vaskemiddeldestabiliserede lipidvesikler og absorption af vaske- eller rengøringsmiddelet eller vaske- og rengøringsmidlerne på polystyrenperler19 eller fjernelse af vaske- og rengøringsmidler ved dialyse eller størrelsesudelukkelseskromatografi20. Organiske opløsningsmidler er blevet brugt til at danne lipidvesikler, for eksempel via dannelsen af olie-vand-interfaser21, men størstedelen af integrerede membranproteiner inaktiveres, når de udsættes for sådanne opløsningsmidler.
I vores laboratorium rekonstituerer vi for det meste membranproteiner ved hjælp af vaskemiddelabsorptionsmetoden for at danne store unilamellære vesikler (LUV’er)19. Denne metode tillader samrekonstituering af flere membranproteiner og indkapsling i vesikellumen af enzymer, metabolitter og prober22,23. De membranproteinholdige LUV’er kan omdannes til gigantiske unilamellære vesikler (GUV’er) med/uden indkapsling af vandopløselige komponenter ved hjælp af enten elektrodannelse24 eller gelassisteret hævelse25 og specifikke betingelser for at bevare membranproteinernes integritet26.
Denne artikel præsenterer en protokol til rekonstituering i LUV’er af et metabolisk netværk uden for ligevægt, der regenererer ATP gennem nedbrydning af L-arginin til L-ornitin27. Dannelsen af ATP er koblet til produktionen af glycerol-3-phosphat (G3P), en vigtig byggesten for fosfolipidsyntese22,28. Den metaboliske vej består af to integrerede membranproteiner, en arginin/ornitin (ArcD) og en G3P/Pi-antiporter (GlpT). Derudover kræves tre opløselige enzymer (ArcA, ArcB, ArcC) til genanvendelse af ATP, og GlpK bruges til at omdanne glycerol til glycerol 3-phosphat ved hjælp af ATP fra nedbrydningen af L-arginin, se figur 1 for en skematisk oversigt over vejen. Denne protokol repræsenterer et godt udgangspunkt for den fremtidige konstruktion af endnu mere komplekse reaktionsnetværk – til syntese af lipider eller proteiner eller deling af celler. Vesiklernes lipidsammensætning understøtter aktiviteten af en lang række integrerede membranproteiner og er optimeret til transport af forskellige molekyler ind i eller ud af vesiklerne 27,29,30.
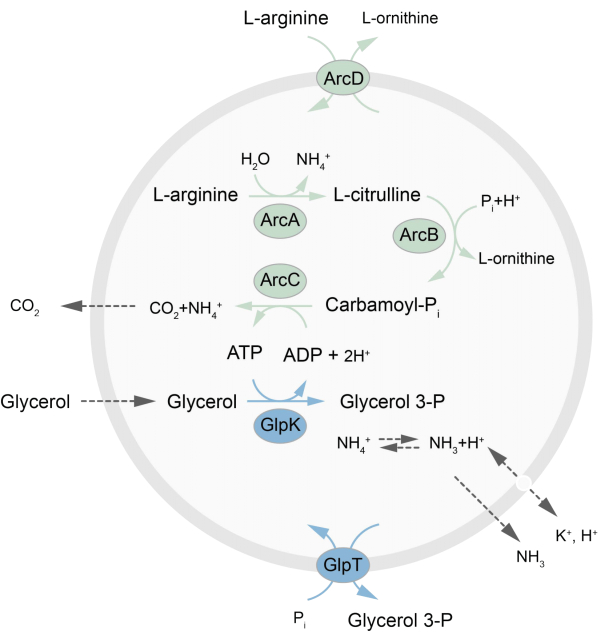
Figur 1: Oversigt over vejen for ATP-produktion og glycerol-3-phosphatsyntese og -udskillelse, klik her for at se en større version af denne figur.
Kort sagt tilsættes rensede membranproteiner (opløst i dodecyl-β-D-maltosid, DDM) til præformede lipidvesikler, der er blevet destabiliseret med Triton X-100, hvilket tillader indsættelse af proteinerne i membranen. Vaskemiddelmolekylerne fjernes efterfølgende (langsomt) ved tilsætning af aktiverede polystyrenperler, hvilket resulterer i dannelsen af godt forseglede proteoliposomer. Opløselige komponenter kan derefter tilsættes vesiklerne og indkapsles via fryse-optøningscyklusser, som fanger molekylerne i membranfusionsprocessen. De opnåede vesikler er meget heterogene og mange er multilamellære. De ekstruderes derefter gennem et polycarbonatfilter med en porestørrelse på 400, 200 eller 100 nm, hvilket giver mere ensartede vesikler; Jo mindre porestørrelsen er, jo mere homogene og unilamellære er vesiklerne, men til prisen for et mindre indre volumen. Ikke-inkorporerede proteiner og små molekyler fjernes fra den eksterne opløsning ved størrelsesudelukkelseskromatografi. ProteoGUL’erne kan omdannes til vesikler i mikrometerstørrelse ved gelassisteret hævelse, og disse proteoGUL’er opsamles derefter og fanges i en mikrofluidisk chip til mikroskopisk karakterisering og manipulation. Figur 2 viser en skematisk oversigt over den fulde protokol.
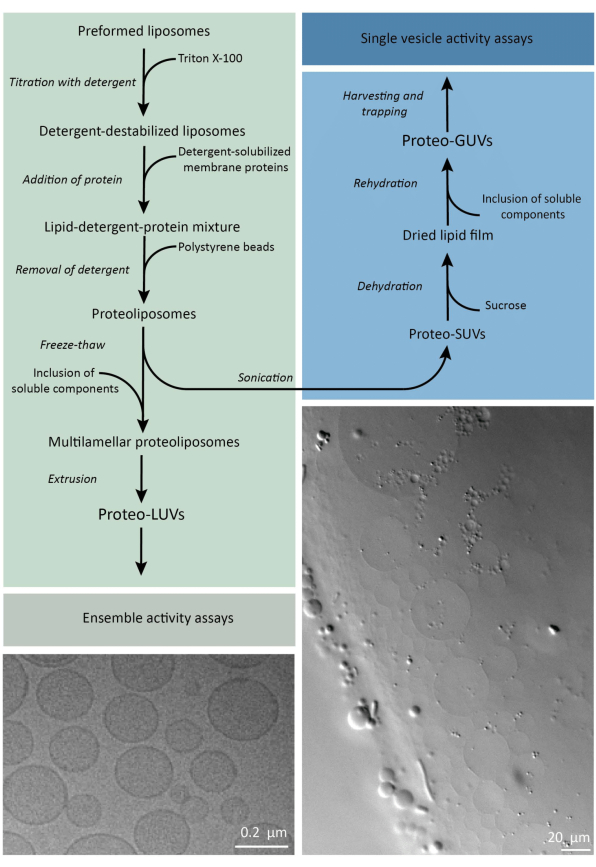
Figur 2: Oversigt over protokollen for rekonstituering af membranproteiner og indkapsling af enzymer og vandopløselige komponenter i lipidvesikler af submikrometer (LV’er) og mikrometerstørrelse (GUV’er).
Rekonstitutions- og indkapslingsprotokollerne fungerer godt, og proteinernes funktionalitet bevares, men proteoGUL’erne og proteoGUV’erne er heterogene i størrelse. Mikrofluidiske tilgange 31,32 tillader dannelse af mikrometerstore vesikler, der er mere homogene i størrelse, men funktionel rekonstitution af membranproteiner er generelt ikke mulig, fordi resterende opløsningsmiddel i dobbeltlaget inaktiverer proteinerne. ProteoUV’erne varierer i størrelse fra 100 til 400 nm, og ved lave koncentrationer af enzymer kan indkapslingen føre til vesikler med ufuldstændige metaboliske veje (stokastiske effekter; se figur 3). LÚV’er er ideelle til at konstruere specifikke metaboliske moduler, som vist her til produktion af ATP og byggesten som G3P. Sådanne proteoGUL’er kan potentielt indkapsles i GUV’er og tjene som organellignende rum for værtsvesiklerne.
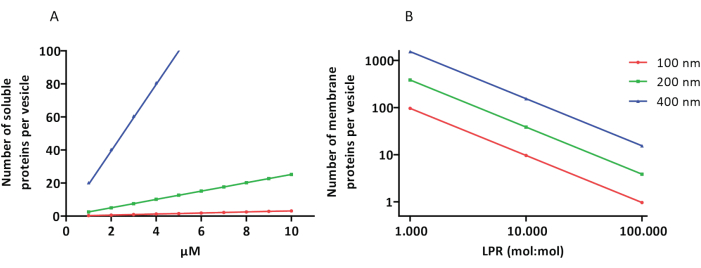
Figur 3: Antal molekyler pr. vesikel med en diameter på 100, 200 eller 400 nm. (A) Når de indkapslede proteiner (enzymer, prober) er i området 1-10 μM. (B) Rekonstitutionen sker ved 1 til 1.000, 1 til 10.000 og 1 til 100.000 membranproteiner pr. lipid (mol/mol). Vi antager, at molekyler er indkapslet i de angivne koncentrationer og inkorporeret i membranen ved disse protein-til-lipid-forhold. For nogle enzymer har vi set, at de binder sig til membraner, hvilket kan øge deres tilsyneladende koncentration i vesiklerne. Forkortelse: LPR = Lipid-Protein-Ratio Klik her for at se en større version af denne figur.
