Området bottom-up syntetisk biologi fokuserar på att konstruera (minimala) celler 1,2 och metaboliska bioreaktorer för bioteknologiska 3,4 eller biomedicinska ändamål 5,6,7,8. Konstruktionen av syntetiska celler ger en unik plattform som gör det möjligt för forskare att studera (membran)proteiner under väldefinierade förhållanden som efterliknar dem i naturliga miljöer, vilket möjliggör upptäckt av emergenta egenskaper och dolda biokemiska funktioner hos proteiner och reaktionsnätverk9. Som ett mellansteg mot en autonomt fungerande syntetisk cell utvecklas moduler som fångar upp väsentliga egenskaper hos levande celler såsom metabolisk energibevarande, protein- och lipidsyntes och homeostas. Sådana moduler förbättrar inte bara vår förståelse av livet utan har också potentiella tillämpningar inom områdena medicin8 och bioteknik10.
Transmembranproteiner är kärnan i praktiskt taget alla metaboliska nätverk eftersom de transporterar molekyler in eller ut ur cellen, signalerar och svarar på miljöns kvalitet och spelar många biosyntetiska roller. Således kräver konstruktionen av metaboliska moduler i syntetiska celler i de flesta fall rekonstituering av integrerade och/eller perifera membranproteiner till ett membrandubbelskikt som består av specifika lipider och hög integritet (låg permeabilitet). Hanteringen av dessa membranproteiner är utmanande och kräver specifik kunskap och experimentella färdigheter.
Flera metoder har utvecklats för att rekonstruera membranproteiner i fosfolipidvesiklar, oftast i syfte att studera funktionen11,12, reglering13, kinetiska egenskaper14,15, lipidberoende15,16 och/eller stabilitet17 hos ett specifikt protein. Dessa metoder innefattar snabb utspädning av tvättmedelslösligt protein till vattenhaltigt medium i närvaro av lipider18, avlägsnande av tvättmedel genom inkubation av tvättmedelslösligt protein med detergentdestabiliserade lipidvesiklar och absorption av tvättmedlet eller tvättmedlen på polystyrenpärlor19, eller avlägsnande av rengöringsmedel genom dialys eller storleksuteslutningskromatografi20. Organiska lösningsmedel har använts för att bilda lipidvesiklar, till exempel genom bildandet av olje-vatten-interfaser21, men majoriteten av integrerade membranproteiner inaktiveras när de utsätts för sådana lösningsmedel.
I vårt laboratorium rekonstituerar vi mestadels membranproteiner med tvättmedelsabsorptionsmetoden för att bilda stora unilamellära vesiklar (LUV)19. Denna metod gör det möjligt att samtidigt rekonstituera flera membranproteiner och kapsla in enzymer, metaboliter och sonder22,23 i vesikellumen. De membranproteininnehållande LUV:erna kan omvandlas till jätte-unilamellära vesiklar (GUV) med/utan inkapsling av vattenlösliga komponenter, med hjälp av antingen elektroformation24 eller gelassisterad svullnad25 och specifika förhållanden för att bevara integriteten hos membranproteinerna26.
Denna artikel presenterar ett protokoll för rekonstituering i LUV:er av ett metaboliskt nätverk som är ur jämvikt och som regenererar ATP genom nedbrytning av L-arginin till L-ornitin27. Bildningen av ATP är kopplad till produktionen av glycerol-3-fosfat (G3P), en viktig byggsten för fosfolipidsyntes22,28. Den metaboliska vägen består av två integrerade membranproteiner, en arginin/ornitin (ArcD) och en G3P/Pi-antiporter (GlpT). Dessutom krävs tre lösliga enzymer (ArcA, ArcB, ArcC) för återvinning av ATP, och GlpK används för att omvandla glycerol till glycerol-3-fosfat, med hjälp av ATP från nedbrytningen av L-arginin, se figur 1 för en schematisk översikt över vägen. Detta protokoll representerar en bra utgångspunkt för den framtida konstruktionen av ännu mer komplexa reaktionsnätverk – för syntes av lipider eller proteiner eller delning av celler. Lipidsammansättningen i vesiklarna stöder aktiviteten hos en mängd olika integrerade membranproteiner och har optimerats för transport av olika molekyler in i eller ut ur vesiklarna 27,29,30.
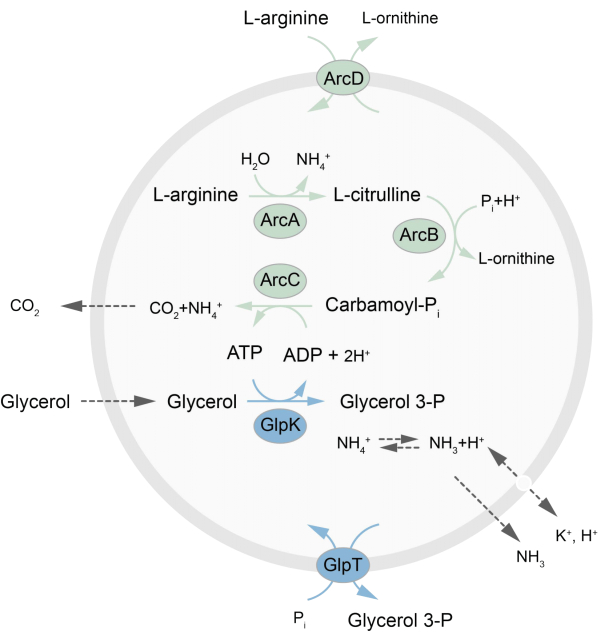
Figur 1: Översikt över vägen för ATP-produktion och syntes och utsöndring av glycerol-3-fosfat. Klicka här för att se en större version av denna figur.
Kort sagt, renade membranproteiner (solubiliserade i dodecyl-β-D-maltosid, DDM) tillsätts till förformade lipidvesiklar som har destabiliserats med Triton X-100, vilket gör det möjligt att föra in proteinerna i membranet. Tvättmedelsmolekylerna avlägsnas därefter (långsamt) genom tillsats av aktiverade polystyrenpärlor, vilket resulterar i bildandet av väl förseglade proteoliposomer. Lösliga komponenter kan sedan tillsättas till vesiklarna och kapslas in via frys-upptiningscykler, vilket fångar molekylerna i processen för membranfusion. De erhållna vesiklarna är mycket heterogena och många är multilamellära vesiklar. De extruderas sedan genom ett polykarbonatfilter med en porstorlek på 400, 200 eller 100 nm, vilket ger mer jämnt dimensionerade vesiklar; Ju mindre porstorlek, desto mer homogena och unilamellära vesiklar men till priset av en mindre inre volym. Icke-inkorporerade proteiner och små molekyler avlägsnas från den externa lösningen genom storleksuteslutningskromatografi. ProteoLUV:erna kan omvandlas till vesiklar i mikrometerstorlek genom gelassisterad svullnad, och dessa proteoGUV:er samlas sedan in och fångas i ett mikrofluidiskt chip för mikroskopisk karakterisering och manipulation. Figur 2 visar en schematisk översikt över det fullständiga protokollet.
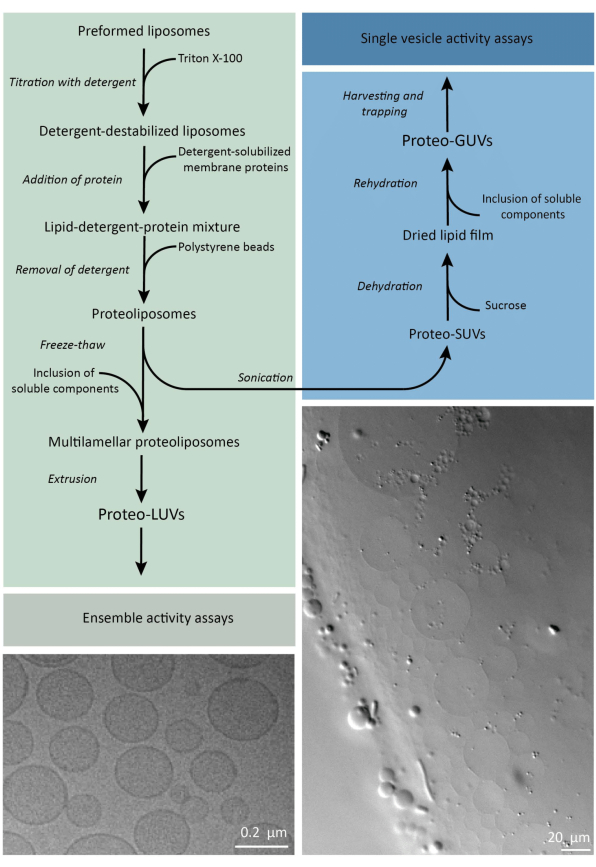
Figur 2: Översikt över protokollet för rekonstituering av membranproteiner och inkapsling av enzymer och vattenlösliga komponenter i lipidvesiklar av submikrometer (LUV) och mikrometerstorlek (GUV). Klicka här för att se en större version av denna figur.
Rekonstituerings- och inkapslingsprotokollen fungerar bra och proteinernas funktionalitet bibehålls, men proteoLUV:erna och proteoGUV:erna är heterogena i storlek. Mikrofluidiska metoder 31,32 tillåter bildandet av mikrometerstora vesiklar som är mer homogena i storlek, men funktionell rekonstitution av membranproteiner är i allmänhet inte möjlig eftersom kvarvarande lösningsmedel i dubbellagret inaktiverar proteinerna. ProteoLUV:erna varierar i storlek från 100 till 400 nm, och vid låga koncentrationer av enzymer kan inkapslingen leda till vesiklar med ofullständiga metaboliska vägar (stokastiska effekter; se figur 3). LUV:er är idealiska för att konstruera specifika metaboliska moduler, som visas här för produktion av ATP och byggstenar som G3P. Sådana proteoLUV:er kan potentiellt kapslas in i GUV:er och fungera som organellliknande fack för värdvesiklarna.
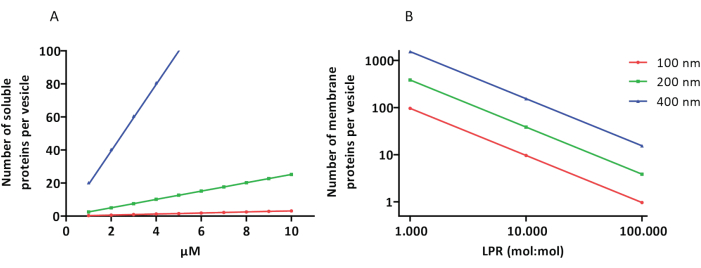
Figur 3: Antal molekyler per vesikel med en diameter på 100, 200 eller 400 nm. (A) När de inkapslade proteinerna (enzymer, sonder) ligger i intervallet 1-10 μM. (B) Beredningen görs vid 1 till 1 000, 1 till 10 000 och 1 till 100 000 membranproteiner per lipid (mol/mol). Vi gör antagandet att molekyler är inkapslade vid de angivna koncentrationerna och inkorporeras i membranet vid dessa protein-till-lipid-förhållanden. För vissa enzymer har vi sett att de binder till membran, vilket kan öka deras skenbara koncentration i vesiklarna. Förkortning: LPR = Lipid-Protein-Ratio Klicka här för att se en större version av denna figur.
