Der Bereich der synthetischen Biologie von unten konzentriert sich auf die Konstruktion von (minimalen) Zellen 1,2 und metabolischen Bioreaktoren für biotechnologische 3,4 oder biomedizinische Zwecke 5,6,7,8. Die Konstruktion synthetischer Zellen bietet eine einzigartige Plattform, die es Forschern ermöglicht, (Membran-)Proteine unter genau definierten Bedingungen zu untersuchen, die denen der natürlichen Umgebung nachahmen, und ermöglicht so die Entdeckung von emergenten Eigenschaften und verborgenen biochemischen Funktionen von Proteinen und Reaktionsnetzwerken9. Als Zwischenschritt hin zu einer autonom funktionierenden synthetischen Zelle werden Module entwickelt, die wesentliche Merkmale lebender Zellen wie metabolische Energieerhaltung, Protein- und Lipidsynthese sowie Homöostase erfassen. Solche Module erweitern nicht nur unser Verständnis des Lebens, sondern haben auch Anwendungsmöglichkeiten in den Bereichen Medizin8 und Biotechnologie10.
Transmembranproteine sind das Herzstück praktisch jedes Stoffwechselnetzwerks, da sie Moleküle in oder aus der Zelle transportieren, Signale senden und auf die Qualität der Umwelt reagieren und zahlreiche biosynthetische Rollen spielen. Daher erfordert das Engineering von Stoffwechselmodulen in synthetischen Zellen in den meisten Fällen die Rekonstitution von integralen und/oder peripheren Membranproteinen zu einer Membrandoppelschicht, die aus spezifischen Lipiden besteht und eine hohe Integrität (geringe Permeabilität) aufweist. Der Umgang mit diesen Membranproteinen ist anspruchsvoll und erfordert spezifisches Wissen und experimentelle Fähigkeiten.
Es wurden mehrere Methoden entwickelt, um Membranproteine in Phospholipid-Vesikeln zu rekonstituieren, meist mit dem Ziel, die Funktion11,12, die Regulation13, die kinetischen Eigenschaften14,15, die Lipidabhängigkeit15,16 und/oder die Stabilität17 eines spezifischen Proteins zu untersuchen. Diese Verfahren umfassen die rasche Verdünnung von durch Detergenzien gelöstem Protein in wässrigen Medien in Gegenwart von Lipiden18, die Entfernung von Detergenzien durch Inkubation von durch Detergenzien gelöstes Protein mit detergenzient destabilisierten Lipidvesikeln und die Absorption des Detergens bzw. der Detergenzien auf Polystyrolkügelchen19 oder das Entfernen von Detergenzien durch Dialyse oder Größenausschlusschromatographie20. Organische Lösungsmittel wurden verwendet, um Lipidvesikel zu bilden, z. B. durch die Bildung von Öl-Wasser-Interphasen21, aber die Mehrzahl der integralen Membranproteine wird inaktiviert, wenn sie solchen Lösungsmitteln ausgesetzt werden.
In unserem Labor rekonstituieren wir hauptsächlich Membranproteine mit der Detergenz-Absorptionsmethode zu großen unilamellären Vesikeln (LUVs)19. Dieses Verfahren ermöglicht die Co-Rekonstitution mehrerer Membranproteine und die Verkapselung von Enzymen, Metaboliten und Sonden im Vesikellumen22,23. Die Membranprotein-haltigen LUVs können in riesen-unilamelläre Vesikel (GUVs) mit/ohne Verkapselung wasserlöslicher Komponenten umgewandelt werden, wobei entweder die Elektroformation24 oder die gelgestützte Quellung25 und spezifische Bedingungen zur Erhaltung der Integrität der Membranproteine26 verwendet werden.
In dieser Arbeit wird ein Protokoll für die Rekonstitution eines außerhalb des Gleichgewichts befindlichen metabolischen Netzwerks in LUVs vorgestellt, das ATP durch den Abbau von L-Arginin in L-Ornithin regeneriert27. Die Bildung von ATP ist an die Produktion von Glycerin-3-phosphat (G3P) gekoppelt, einem wichtigen Baustein für die Phospholipidsynthese22,28. Der Stoffwechselweg besteht aus zwei integralen Membranproteinen, einem Arginin/Ornithin (ArcD) und einem G3P/Pi-Antiporter (GlpT). Darüber hinaus werden drei lösliche Enzyme (ArcA, ArcB, ArcC) für das Recycling von ATP benötigt, und GlpK wird verwendet, um Glycerin in Glycerin-3-phosphat umzuwandeln, wobei das ATP aus dem Abbau von L-Arginin verwendet wird, siehe Abbildung 1 für einen schematischen Überblick über den Weg. Dieses Protokoll stellt einen guten Ausgangspunkt für den zukünftigen Aufbau noch komplexerer Reaktionsnetzwerke dar – für die Synthese von Lipiden oder Proteinen oder die Teilung von Zellen. Die Lipidzusammensetzung der Vesikel unterstützt die Aktivität einer Vielzahl von integralen Membranproteinen und wurde für den Transport verschiedener Moleküle in oder aus den Vesikeln optimiert 27,29,30.
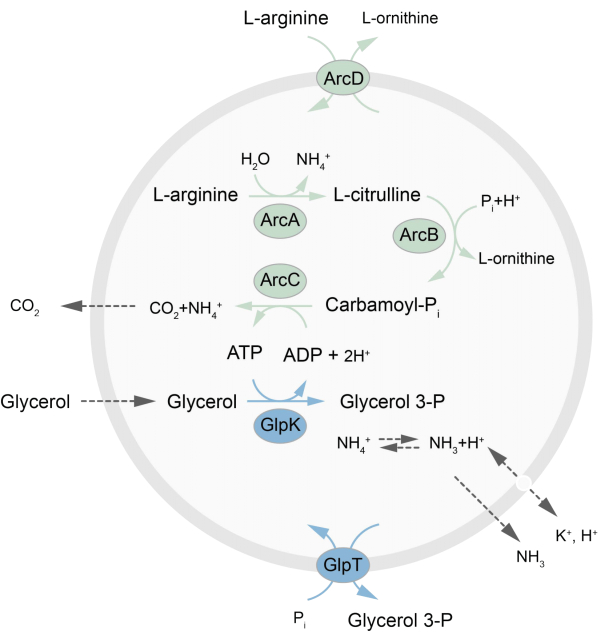
Abbildung 1: Überblick über den Weg der ATP-Produktion und der Glycerin-3-phosphat-Synthese und -Ausscheidung. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
Kurz gesagt, gereinigte Membranproteine (solubilisiert in Dodecyl-β-D-Maltosid, DDM) werden zu vorgeformten Lipidvesikeln hinzugefügt, die mit Triton X-100 destabilisiert wurden, was die Insertion der Proteine in die Membran ermöglicht. Die Detergensmoleküle werden anschließend (langsam) durch die Zugabe von aktivierten Polystyrolkügelchen entfernt, was zur Bildung von gut verschlossenen Proteoliposomen führt. Lösliche Bestandteile können dann zu den Vesikeln hinzugefügt und über Gefrier-Tau-Zyklen verkapselt werden, wodurch die Moleküle im Prozess der Membranfusion eingefangen werden. Die gewonnenen Vesikel sind sehr heterogen und viele sind multilamell. Sie werden dann durch einen Polycarbonatfilter mit einer Porengröße von 400, 200 oder 100 nm extrudiert, was zu gleichmäßiger großen Vesikeln führt; Je kleiner die Porengröße, desto homogener und unilamellär sind die Vesikel, jedoch zum Preis eines kleineren inneren Volumens. Nicht eingebaute Proteine und kleine Moleküle werden durch Größenausschlusschromatographie aus der externen Lösung entfernt. Die proteoLUVs können durch gelgestütztes Aufquellen in mikrometergroße Vesikel umgewandelt werden, und diese proteoGUVs werden dann gesammelt und in einem mikrofluidischen Chip zur mikroskopischen Charakterisierung und Manipulation gefangen. Abbildung 2 zeigt einen schematischen Überblick über das gesamte Protokoll.
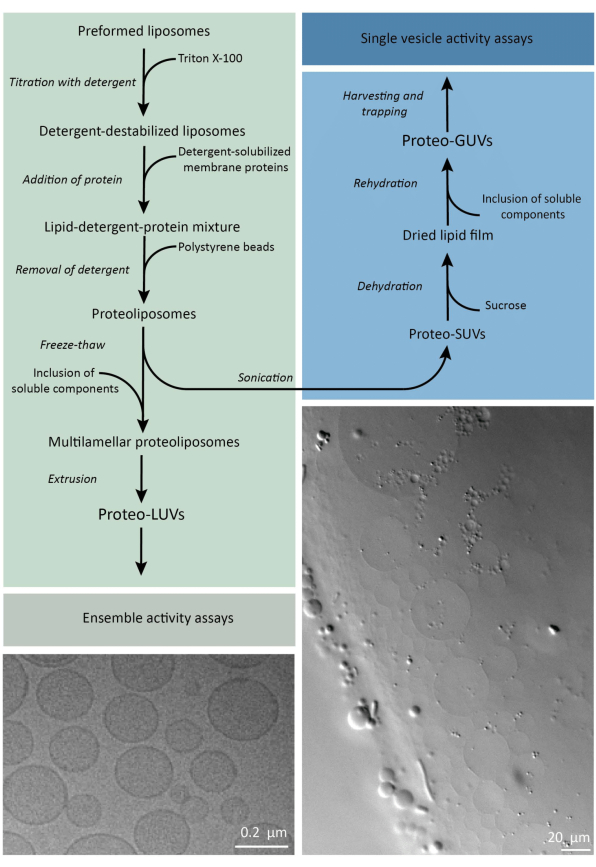
Abbildung 2: Überblick über das Protokoll zur Rekonstitution von Membranproteinen und zur Verkapselung von Enzymen und wasserlöslichen Komponenten in Lipidvesikeln mit Submikrometer- (LUVs) und Mikrometergröße (GUVs). Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
Die Rekonstitutions- und Verkapselungsprotokolle funktionieren gut und die Funktionalität der Proteine bleibt erhalten, aber die proteoLUVs und proteoGUVs sind heterogen groß. Mikrofluidische Ansätze 31,32 ermöglichen die Bildung von mikrometergroßen Vesikel, die in ihrer Größe homogener sind, aber eine funktionelle Rekonstitution von Membranproteinen ist im Allgemeinen nicht möglich, da das restliche Lösungsmittel in der Doppelschicht die Proteine inaktiviert. Die ProteoLUVs haben eine Größe von 100 bis 400 nm, und bei niedrigen Enzymkonzentrationen kann die Verkapselung zu Vesikeln mit unvollständigen Stoffwechselwegen führen (stochastische Effekte; siehe Abbildung 3). LUVs sind ideal für den Aufbau spezifischer Stoffwechselmodule, wie hier gezeigt, für die Produktion von ATP und Bausteinen wie G3P. Solche proteoLUVs können möglicherweise in GUVs eingekapselt werden und als organellenähnliche Kompartimente für die Wirtsvesikel dienen.
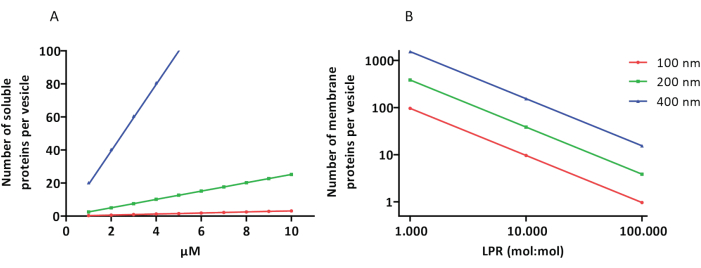
Abbildung 3: Anzahl der Moleküle pro Vesikel mit einem Durchmesser von 100, 200 oder 400 nm. (A) Wenn die verkapselten Proteine (Enzyme, Sonden) im Bereich von 1-10 μM liegen. (B) Die Rekonstitution erfolgt bei 1 bis 1.000, 1 bis 10.000 und 1 bis 100.000 Membranproteinen pro Lipid (mol/mol). Wir gehen davon aus, dass Moleküle in den angegebenen Konzentrationen verkapselt und bei diesen Protein-Lipid-Verhältnissen in die Membran eingebaut werden. Bei einigen Enzymen haben wir gesehen, dass sie an Membranen binden, was ihre scheinbare Konzentration in den Vesikeln erhöhen kann. Abkürzung: LPR = Lipid-Protein-Ratio Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
