नीचे-ऊपर सिंथेटिक जीव विज्ञान के क्षेत्र (न्यूनतम) कोशिकाओं 1,2 और जैव प्रौद्योगिकी 3,4 या जैव चिकित्सा प्रयोजनों 5,6,7,8 के लिए चयापचय bioreactors के निर्माण पर केंद्रित है. सिंथेटिक कोशिकाओं का निर्माण एक अनूठा मंच प्रदान करता है जो शोधकर्ताओं को देशी वातावरण की नकल करने वाली अच्छी तरह से परिभाषित स्थितियों में प्रोटीन का अध्ययन करने की अनुमति देता है, जिससे प्रोटीन औरप्रतिक्रिया नेटवर्क 9 के आकस्मिक गुणों और छुपा जैव रासायनिक कार्यों की खोज को सक्षम किया जा सकता है। एक स्वायत्त रूप से काम करने वाले सिंथेटिक सेल की दिशा में एक मध्यवर्ती कदम के रूप में, मॉड्यूल विकसित किए जाते हैं जो चयापचय ऊर्जा संरक्षण, प्रोटीन और लिपिड संश्लेषण, और होमियोस्टेसिस जैसे जीवित कोशिकाओं की आवश्यक विशेषताओं को पकड़ते हैं। इस तरह के मॉड्यूल न केवल जीवन की हमारी समझ को बढ़ाने के लिए लेकिन यह भी चिकित्सा8 और जैव प्रौद्योगिकी10 के क्षेत्र में संभावित अनुप्रयोगों है.
ट्रांसमेम्ब्रेन प्रोटीन वस्तुतः किसी भी चयापचय नेटवर्क के केंद्र में होते हैं क्योंकि वे अणुओं को कोशिका के अंदर या बाहर ले जाते हैं, संकेत देते हैं, और पर्यावरण की गुणवत्ता का जवाब देते हैं, और कई बायोसिंथेटिक भूमिकाएं निभाते हैं। इस प्रकार, सिंथेटिक कोशिकाओं में चयापचय मॉड्यूल की इंजीनियरिंग के लिए ज्यादातर मामलों में विशिष्ट लिपिड और उच्च अखंडता (कम पारगम्यता) से बना झिल्ली बाईलेयर में अभिन्न और / या परिधीय झिल्ली प्रोटीन के पुनर्गठन की आवश्यकता होती है। इन झिल्ली प्रोटीन की हैंडलिंग चुनौतीपूर्ण है और विशिष्ट ज्ञान और प्रयोगात्मक कौशल की आवश्यकता है.
फॉस्फोलिपिड पुटिकाओं के भीतर झिल्ली प्रोटीन का पुनर्गठन करने के लिए कई तरीकों का विकास किया गया है, अक्सर समारोह11,12, विनियमन13, गतिज गुण14,15, लिपिड निर्भरता15,16, और/या स्थिरता17 एक विशिष्ट प्रोटीन के अध्ययन के उद्देश्य से। इन विधियों लिपिड18 की उपस्थिति में जलीय मीडिया में डिटर्जेंट घुलनशील प्रोटीन के तेजी से कमजोर पड़ने शामिल, डिटर्जेंट-अस्थिर लिपिड पुटिकाओं और डिटर्जेंट के अवशोषण के साथ डिटर्जेंट-घुलनशील प्रोटीन सेते द्वारा डिटर्जेंट को हटाने(रों) polystyrene मोती19 पर, या डायलिसिस या आकार बहिष्करण क्रोमैटोग्राफी द्वारा डिटर्जेंट को हटाने20. कार्बनिक सॉल्वैंट्स लिपिड पुटिकाओं के रूप में इस्तेमाल किया गया है, उदाहरण के लिए, तेल-पानी interphases21 के गठन के माध्यम से, लेकिन अभिन्न झिल्ली प्रोटीन के बहुमत निष्क्रिय कर रहे हैं जब इस तरह के सॉल्वैंट्स के संपर्क में हैं.
हमारी प्रयोगशाला में, हम ज्यादातर बड़े-यूनिलामेलर पुटिकाओं (एलयूवी)19बनाने के लिए डिटर्जेंट-अवशोषण विधि द्वारा झिल्ली प्रोटीन का पुनर्गठन करते हैं। यह विधि कई झिल्ली प्रोटीन के सह-पुनर्गठन और एंजाइमों, चयापचयों और जांच22,23 के पुटिका लुमेन में एनकैप्सुलेशन की अनुमति देती है। झिल्ली प्रोटीन युक्त LUVs विशाल unilamellar vesicles (GUVs) के साथ / पानी में घुलनशील घटकों के encapsulation के बिना, या तो electroformation24 या जेल सहायता प्राप्त सूजन25 और झिल्ली प्रोटीन26 की अखंडता को संरक्षित करने के लिए विशिष्ट शर्तों का उपयोग कर में परिवर्तित किया जा सकता है.
यह पत्र एक आउट-ऑफ-इक्विलिब्रियम मेटाबोलिक नेटवर्क के एलयूवी में पुनर्गठन के लिए एक प्रोटोकॉल प्रस्तुत करता है जो एल-आर्जिनिन के टूटने के माध्यम से एटीपी को एल-ऑर्निथिन27 में पुन: उत्पन्न करता है। एटीपी का गठन ग्लिसरॉल-3-फॉस्फेट (जी 3 पी) के उत्पादन के लिए युग्मित है, फॉस्फोलिपिड संश्लेषण 22,28के लिए एक महत्वपूर्ण बिल्डिंग ब्लॉक। चयापचय मार्ग में दो अभिन्न झिल्ली प्रोटीन, एक आर्जिनिन / ऑर्निथिन (एआरसीडी) और एक जी 3 पी / पीआई एंटीपोर्टर (जीएलपीटी) होते हैं। इसके अलावा, एटीपी के पुनर्चक्रण के लिए तीन घुलनशील एंजाइम (ArcA, ArcB, ArcC) की आवश्यकता होती है, और GlpK का उपयोग ग्लिसरॉल को ग्लिसरॉल 3-फॉस्फेट में बदलने के लिए किया जाता है, L-arginine के टूटने से ATP का उपयोग करके, मार्ग के योजनाबद्ध अवलोकन के लिए चित्र 1 देखें। यह प्रोटोकॉल लिपिड या प्रोटीन के संश्लेषण या कोशिकाओं के विभाजन के लिए और भी जटिल प्रतिक्रिया नेटवर्क के भविष्य के निर्माण के लिए एक अच्छा प्रारंभिक बिंदु का प्रतिनिधित्व करता है। पुटिकाओं की लिपिड संरचना अभिन्न झिल्ली प्रोटीन की एक विस्तृत विविधता की गतिविधि का समर्थन करती है और पुटिकाओं 27,29,30में या बाहर विविध अणुओं के परिवहन के लिए अनुकूलित किया गया है।
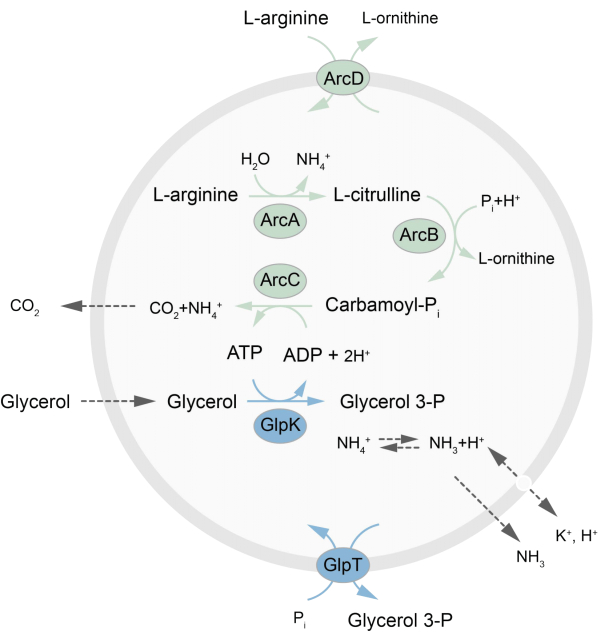
चित्रा 1: एटीपी उत्पादन और ग्लिसरॉल 3-फॉस्फेट संश्लेषण और उत्सर्जन के लिए मार्ग का अवलोकन। कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें।
संक्षेप में, शुद्ध झिल्ली प्रोटीन (डोडेसिल-β-डी-माल्टोसाइड, डीडीएम में घुलनशील) को पूर्वनिर्मित लिपिड पुटिकाओं में जोड़ा जाता है जिन्हें ट्राइटन एक्स -100 के साथ अस्थिर किया गया है, जो झिल्ली में प्रोटीन के सम्मिलन की अनुमति देता है। डिटर्जेंट अणुओं को बाद में (धीरे-धीरे) सक्रिय पॉलीस्टायर्न मोतियों के अतिरिक्त हटा दिया जाता है, जिसके परिणामस्वरूप अच्छी तरह से सील प्रोटीओलिपोसोम का निर्माण होता है। घुलनशील घटकों को तब पुटिकाओं में जोड़ा जा सकता है और फ्रीज-पिघलना चक्रों के माध्यम से समझाया जा सकता है, जो झिल्ली संलयन की प्रक्रिया में अणुओं को फँसाता है। प्राप्त पुटिकाएं अत्यधिक विषम होती हैं और कई मल्टीलामेलर होती हैं। फिर उन्हें 400, 200, या 100 एनएम के छिद्र आकार के साथ एक पॉली कार्बोनेट फिल्टर के माध्यम से निकाला जाता है, जो अधिक समान आकार के पुटिकाओं का उत्पादन करता है; छिद्र का आकार जितना छोटा होता है, पुटिकाओं का आकार उतना ही अधिक सजातीय और एकमिलामेलर होता है, लेकिन एक छोटे आंतरिक आयतन की कीमत पर। गैर-निगमित प्रोटीन और छोटे अणुओं को आकार-बहिष्करण क्रोमैटोग्राफी द्वारा बाहरी समाधान से हटा दिया जाता है। प्रोटीओएलयूवी को जेल-असिस्टेड सूजन द्वारा माइक्रोमीटर आकार के पुटिकाओं में परिवर्तित किया जा सकता है, और इन प्रोटीओजीयूवी को तब सूक्ष्म लक्षण वर्णन और हेरफेर के लिए माइक्रोफ्लुइडिक चिप में एकत्र और फंसाया जाता है। चित्रा 2 पूर्ण प्रोटोकॉल का एक योजनाबद्ध अवलोकन दिखाता है।
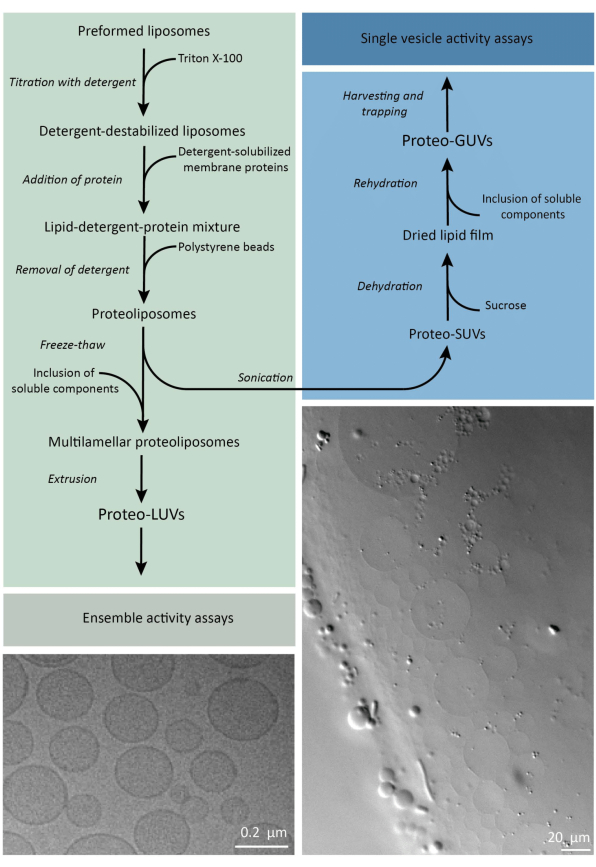
चित्रा 2: झिल्ली प्रोटीन के पुनर्गठन और उप-माइक्रोमीटर (एलयूवी) और माइक्रोमीटर आकार (जीयूवी) के लिपिड पुटिकाओं में एंजाइमों और पानी में घुलनशील घटकों को एनकैप्सुलेट करने के लिए प्रोटोकॉल का अवलोकन। कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें।
पुनर्गठन और एनकैप्सुलेशन प्रोटोकॉल अच्छी तरह से काम करते हैं और प्रोटीन की कार्यक्षमता को बरकरार रखा जाता है, लेकिन प्रोटीओएलयूवी और प्रोटीओजीयूवी आकार में विषम हैं। माइक्रोफ्लुइडिक दृष्टिकोण31,32 माइक्रोमीटर आकार के पुटिकाओं के गठन की अनुमति देते हैं जो आकार में अधिक सजातीय होते हैं, लेकिन झिल्ली प्रोटीन का कार्यात्मक पुनर्गठन आमतौर पर संभव नहीं होता है क्योंकि बाइलेयर में अवशिष्ट विलायक प्रोटीन को निष्क्रिय करता है। प्रोटीओएलयूवी आकार में 100 से 400 एनएम तक होते हैं, और एंजाइमों की कम सांद्रता पर, एनकैप्सुलेशन अपूर्ण चयापचय मार्गों (स्टोकेस्टिक प्रभाव; चित्र 3 देखें) के साथ पुटिकाओं को जन्म दे सकता है। LUVs विशिष्ट चयापचय मॉड्यूल के निर्माण के लिए आदर्श हैं, जैसा कि एटीपी के उत्पादन और G3P जैसे बिल्डिंग ब्लॉक्स के लिए यहां दिखाया गया है। इस तरह के प्रोटीओएलयूवी को संभावित रूप से जीयूवी में समझाया जा सकता है और मेजबान पुटिकाओं के लिए ऑर्गेनेल जैसे डिब्बों के रूप में काम किया जा सकता है।
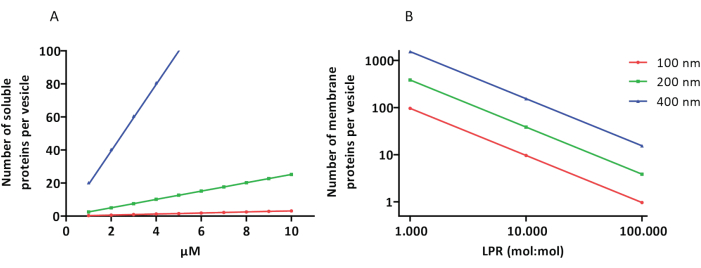
चित्रा 3: 100, 200, या 400 एनएम के व्यास के साथ प्रति पुटिका अणुओं की संख्या। (ए) जब समझाया प्रोटीन (एंजाइम, जांच) 1-10 माइक्रोन की सीमा में हैं। (बी) पुनर्गठन 1 से 1,000, 1 से 10,000, और 1 से 100,000 झिल्ली प्रोटीन प्रति लिपिड (मोल / मोल) पर किया जाता है। हम यह धारणा बनाते हैं कि अणुओं को संकेतित सांद्रता पर समझाया जाता है और इन प्रोटीन-से-लिपिड अनुपात में झिल्ली में शामिल किया जाता है। कुछ एंजाइमों के लिए, हमने देखा है कि वे झिल्ली से बंधते हैं, जो पुटिकाओं में उनकी स्पष्ट एकाग्रता को बढ़ा सकते हैं। संक्षिप्तीकरण: LPR = लिपिड-प्रोटीन-अनुपात कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें।
