The cell samples were treated as described in the protocol and flow cytometry detection was done. Normal cells cannot be stained with Annexin V and 7-AAD (Annexin V-/7-AAD-). In the early stages of pyroptosis, apoptosis, and necroptosis, PS was exposed and bound to Annexin V, but the cell membrane was still intact and excluded 7-AAD from the extracellular space (Annexin V+/7-AAD-). In the later stages, the cell membrane loses its integrity, cells are simultaneously stained by Annexin V and 7-AAD, showing double positive results (Annexin V+/7-AAD+). The necrotic cells were only stained with 7-AAD (Annexin V-/7-AAD+). Figure 1 shows that after LPS/ATP stimulation, the cells in the Q2 region increased significantly, indicating cell death.
We use SEM to capture the plasma membrane surface, directly observing the characteristics of membrane from a single cell. Figure 2A shows the control cells, Figure 2B-D show the cells after LPS/ATP stimulation. We observed cells with characteristics of pyroptosis, including membrane blebbing and rupture membrane (Figure 2B), cells with apoptotic characteristics, including membrane blebbing and cell shrinkage (Figure 2C), and cells with necroptosis characteristics, including cell swelling and rupture membrane (Figure 2D).
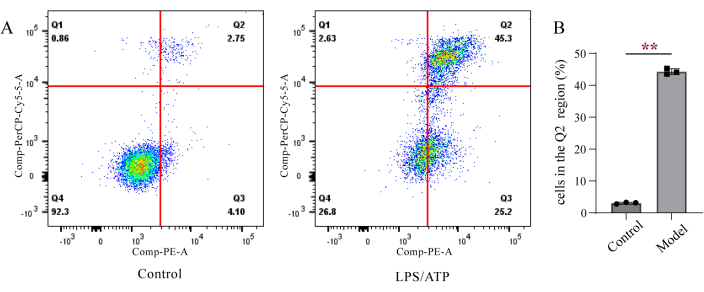
Figure 1: Detection of Annexin V/7-AAD double staining using flow cytometry. (A) Annexin V negative and 7-AAD negative staining (Q4), Annexin V positive and 7-AAD negative double staining (Q3), Annexin V positive and 7-AAD positive staining (Q2), and Annexin V negative and 7-AAD positive staining (Q1). (B) Quantitative data for (A). Data are presented as mean ± SD (n=3), unpaired t test, **p < 0.01. Please click here to view a larger version of this figure.
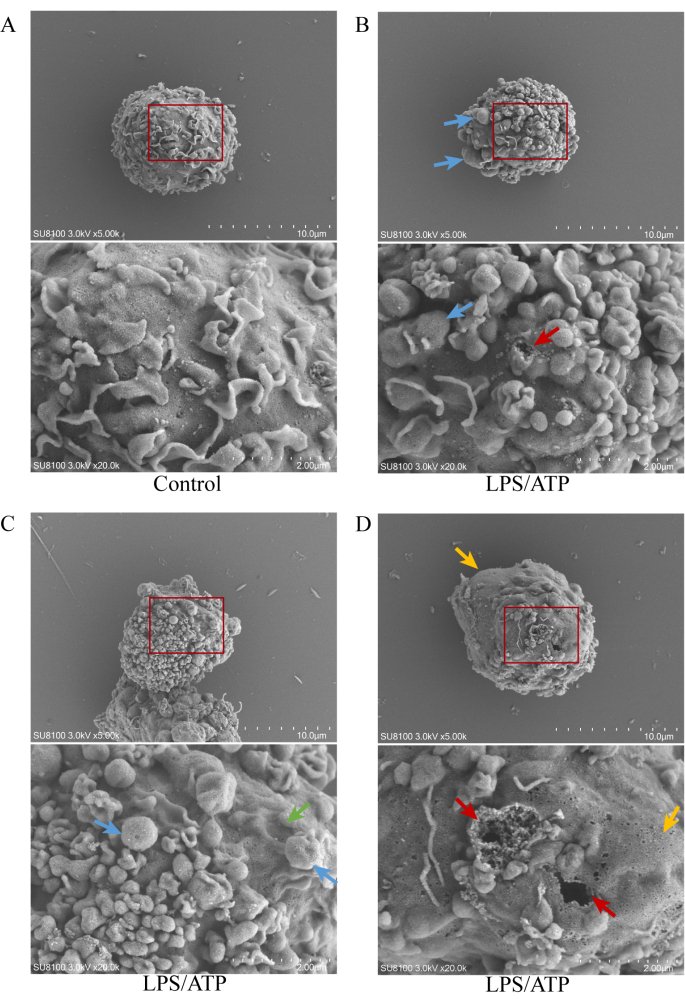
Figure 2: Scanning electron microscopy images of PMA-differentiated THP-1 macrophages. (A) Control group, 5,000x/20,000x magnification. (B, C, D) LPS/ATP group show membrane blebbing, rupture of membrane (↑), cell swelling (↑), and cell shrinkage (↑) at 5,000x/20,000x magnification. Please click here to view a larger version of this figure.
| 0.25% pancreatic enzyme solution (excluding EDTA) | BOSTER Biological Technology co.ltd | PYG0068 | |
| 5 mL Polystyrene Round-Bottom Tube | CORNING | 352235 | |
| 5 mL centrifuge tube | Labgic Technology Co., Ltd. | BS-50-M | |
| 6-well plate | Sorfa Life Science Research Co.,Ltd | 220100 | |
| Annexin V-PE/7-AAD apoptosis analysis kit | Absin (Shanghai) Biological Technology co.ltd | abs50007 | Annexin V-PE, 7-AAD, 5×Binding buffer, Apoptosis Positive Control Solution |
| celculture CO2 incubator | Esco (Shanghai) Enterprise Development Co., Ltd. | N/A | |
| cell culture dish, 100 mm | Sorfa Life Science Research Co.,Ltd | 230301 | |
| Cellometer K2 Fluorescent Cell Counter | Nexcelom Bioscience LLC | Cellometer K2 | |
| Cellometer SD100 Counting Chambers | Nexcelom Bioscience LLC | CHT4-SD100-002 | |
| centrifuge machine | Hunan Xiangyi Laboratory Instrument Development Co., Ltd | L530 | |
| chromium alum | Guangdong Wengjiang Chemical Reagent Co., Ltd. | PA04354 | |
| cover glasses, 9 mm | Labgic Technology Co., Ltd. | BS-09-RC | |
| critical point dryer | Quorum Technologies | K850 | |
| dimethyl sulfoxide | BOSTER Biological Technology co.ltd | PYG0040 | |
| electron microscope fixative | Servicebio Technology co.ltd | G1102 | 2.5% glutaric dialdehyde, 100 mM phosphorous salts |
| electronic balance | SHIMADZU | ATX124 | |
| ethanol absolute | Chengdu Kelong Chemical Co., Ltd | 2021033102 | |
| flow cytometer | Becton,Dickinson and Company | FACSCanto  |
|
| flow cytometry analysis software | Becton,Dickinson and Company | BD FACSDivaTM Software | |
| gelatin | Guangdong Wengjiang Chemical Reagent Co., Ltd. | PA00256 | |
| High resolution cold field emission scanning electron microscope | TITACHI | Regulus 8100 | |
| human monocytic cell line THP-1 | Procell Life Science&Technology Co.,Ltd. | CL0233 | |
| inverted microscope | Leica Microsystems Co., Ltd | DMi1 | |
| IR Vortex Mixer | VELP Scientifica Srl | ZX4 | |
| lipopolysaccharide | Beijing Solarbio Science & Technology Co.,Ltd. | L8880 | LPS is derived from Escherichia coli 055:B5 |
| Na2ATP | Beijing Solarbio Science & Technology Co.,Ltd. | A8270 | |
| phorbol-12-myristate-13-acetate | Beijing Solarbio Science & Technology Co.,Ltd. | P6741 | |
| phosphate-buffered saline | Servicebio Technology co.ltd | G4202 | |
| Pipette | Eppendorf AG | N/A | |
| pipette tips, 10 μL | Servicebio Technology co.ltd | T-10PL | |
| pipette tips, 1 mL | Servicebio Technology co.ltd | T-1250L | |
| pipette tips, 200 μL | Servicebio Technology co.ltd | T-200L | |
| RPMI-1640 complete culture media | Procell Life Science&Technology Co.,Ltd. | CM0233 | RPMI-1640 + 10% FBS + 0.05mM β-mercaptoethanol + 1% P/S |
| RPMI-1640 culture media | Shanghai BasalMedia Technologies Co., LTD. | K211104 | |
| sheath fluid | BECKMAN COULTER | 8546733 | |
| sputter coater | Cressington Scientific Instruments Ltd | 108 | |
| thermostatic water bath | GUOHUA Electric Appliance CO.,Ltd | HH-1 |
