Secretion of acid is a crucial function of the human stomach. However, to what extent acid secretion can be modeled in organoids is still a matter of debate6,32,33,34. We therefore developed the protocol detailed above to accurately measure acid production in gastric organoids. Notably, we used unstimulated adult stem cell-derived organoids cultured under standard expansion conditions that had been passaged several times, which led to parietal cell loss35. Therefore, the presence of acid-secreting parietal cells and active acid release was not expected in our model system.
For our experiments, we used organoids with diameters between 200 µm and 1,000 µm seeded on glass-bottom dishes. First, we tested two different tip sizes for the microelectrodes-a pH-25 with a tip diameter of 25 µm and a pH-50 with a tip diameter of 50 µm. As shown in Figure 2A, there was no significant difference between measurements obtained with the smaller compared to the larger tips. Interestingly, the baseline pH in the organoids tended to be slightly alkaline at ~pH 8. Using the 25 µm pH-25 tip, we next assessed the variation in luminal pH within different ten organoids maintained on the same culture plate, with three measurements obtained from each organoid (Figure 2B). Within one plate, the luminal pH of individual organoids showed only slight, non-significant variations (8.16 ± 0.12; p ≥0.0445-0.99) (Figure 2B). We also compared intraluminal pH measurements within five different organoid lines with passages ranging from 3 to 16 (Figure 2C). Again, we found consistent pH measurements within each culture but significant variability between organoid lines. However, luminal pH generally remained between pH 7.3 and pH 8.2, within one order of magnitude, and there was no apparent trend for average pH when comparing early to late passage numbers (Figure 2C). We next asked whether the pH of the organoid lumen was directly related to the pH of the organoid expansion media and extracellular matrix (ECM). Comparison between the media, the ECM surrounding the organoids, and the organoid lumen revealed significant differences in pH, with the luminal pH of the organoids lower than that of the ECM, and the ECM pH lower than that of the surrounding organoid expansion media (Figure 2D), suggesting that the luminal pH of the organoids was physiologically relevant rather than directly determined by the culture environment. Across six independent experiments, we measured an average luminal pH that was near neutral at 7.13 ± 0.09.
A motorized micromanipulator was used to obtain pH measurements of the organoid lumen with greater spatial resolution. This approach is relevant if pH or other gradients within the mucus-filled organoid lumen29 need to be recorded. Figure 2E shows two representative series of pH measurements in large organoids (>1,000 µm diameter), demonstrating the entry point into each organoid and ending at a depth of ~800 µm. As the two organoids profiled did not have the same diameter, the measurements are not immediately comparable. Regardless, we show evidence of a slight pH gradient of Δ0.6 between the epithelial surface and the deeper organoid lumen (Figure 2E). To determine the feasibility of performing pH measurements in the organoids over time, which would enable measurements of treatment responses in real time, we recorded the intraluminal pH inside a representative organoid for approximately 20 min and found that the reading remained highly consistent after an initial adaptation period (Figure 2F). To validate the luminal pH measurements with an independent method, we used a pH-sensitive colorimetric dye (methyl red) that we injected into the organoid lumen using a micromanipulator-mounted nanoliter autoinjector (Figure 2G). The yellow coloring of the dye confirmed that the organoid lumen had a pH >6.2, consistent with the microelectrode measurements that showed a near-neutral pH. Overall, these representative results illustrate the feasibility and reproducibility of microelectrode-based pH measurements in organoid cultures.
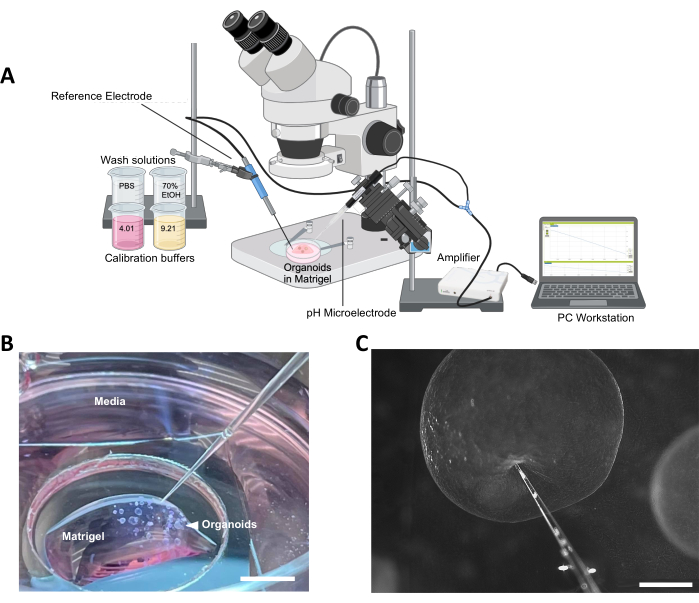
Figure 1: Overview of the method. (A) Schematic diagram of profiling setup. The microelectrode in Figure 1A was taken from the Unisense website.36 (B) Representative image of organoid culture ready for profiling, surrounding ECM, and surrounding media (scale bar = 5 mm). (C) Example of correct microelectrode positioning in preparation for organoid probing (scale bar = 500 µm). Abbreviation: ECM = extracellular matrix. Please click here to view a larger version of this figure.
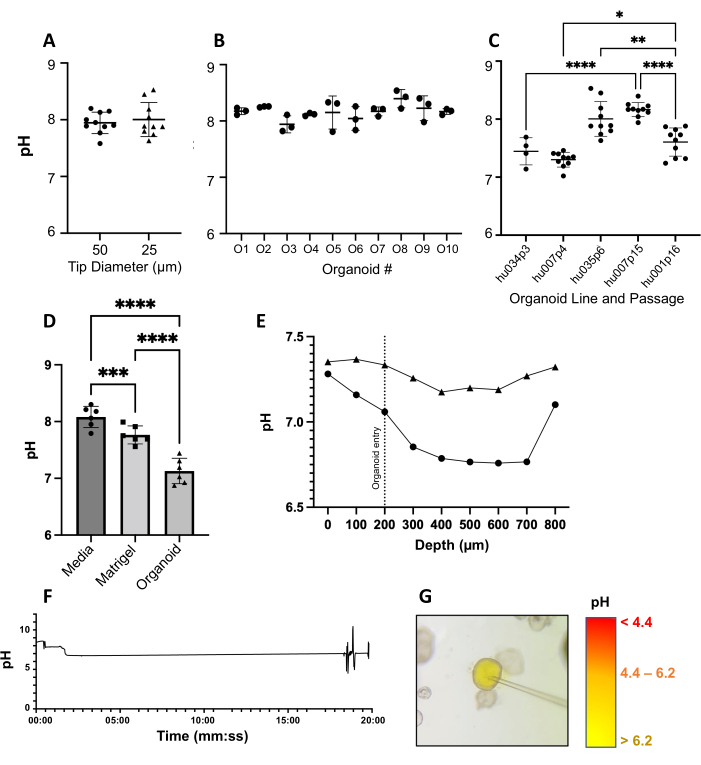
Figure 2: Validation of microsensor profiling in human gastric organoids. (A) Comparison of mid-lumen pH measurements using 25 µm (pH-25) and 50 µm (pH-50) microelectrode tips. Data from 10 individual organoids, each from a single representative experiment. (B) Three replicate pH measurements were obtained for 10 individual organoids. One-way ANOVA with multiple comparisons (p = 0.1229). (C) pH measurements obtained with a pH-25 electrode are consistent within each of five organoid lines analyzed at different passage numbers; 4-10 organoids measured per line based on size and availability. One-way ANOVA with multiple comparisons (p < 0.0001). (D) pH measurements within gastric organoids and the surrounding Matrigel and media (n = 6 independent experiments). Measurements obtained by penetrating gastric epithelial organoids with a pH-25. Each data point is the mean of 10 individual pH measurements within a single organoid. One-way ANOVA (p < 0.0001). (E) Epithelial-to-lumen pH profiles of two representative organoids using a motorized micromanipulator. (F) Stability of pH over time (~19.8 min) in one representative organoid. (G) Microinjection of methyl red pH indicator dye and HCl into the gastric organoid lumen. Please click here to view a larger version of this figure.
| Donor | |||||
| Figure | Line | Passage | Sex | Age | Ethnicity |
| 2A | 35 | 6 | F | 40 | B |
| 2B | 7 | 15 | F | 26 | B |
| 7 | 4 | F | 26 | B | |
| 1 | 16 | F | 45 | C | |
| 34 | 3 | F | 44 | Unknown | |
| 35 | 6 | F | 40 | B | |
| 2C | 7 | 15 | F | 26 | B |
| 2D | 36 | 1 | Unknown | ||
| 7 | 5 | F | 26 | B | |
| 37 | 1 | Unknown | |||
| 31 | 3 | F | 45 | H | |
| 7 | 9 | F | 26 | B | |
| 7 | 4 | F | 26 | B | |
| 2E | 10 | 15 | F | 43 | Unknown |
| 2F | 34 | 3 | F | 33 | C |
| 2G | 7 | 5 | F | 26 | B |
Table 1.
| Human Gastric Organoid Expansion Medium (L-WRN) | |
| L-WRN conditioned medium | 50% |
| Advanced DMEM/F12 | 37% |
| Fetal bovine serum | 10% |
| Penicillin/streptomycin | 1% |
| L-glutamine | 1% |
| Gentamycin | 0.10% |
| Amphotericin B | 0.10% |
| HEPES buffer | 0.40% |
| Y-27632 | 0.10% |
| SB-431542 | 0.10% |
Table 2.
Supplemental Figure S1: Ion transporter expression in human gastric organoids. Relative copy numbers normalized to 18S rRNA. Please click here to download this File.
Supplemental Figure S2: Human gastric organoid with deformed architecture following puncture with the pH microsensor probe. Scale bar = 1 mm. Please click here to download this File.
Supplemental Video S1: Unsuccessful profiling attempt in a gastric organoid culture. The 25 µm pH microelectrode is manually advanced toward the organoid of interest. After a failed attempt due to an angle misjudgment, a different angle is attempted and it is revealed that the organoid lacks the structural integrity to be profiled (spherical structure is not sturdy and deforms upon penetration with the electrode). The user then redirects to attempt a different organoid. Please click here to download this File.
