These protocols were applied to 50 expectorated sputum samples from pwCF presenting for routine care to an outpatient cystic fibrosis clinic at Massachusetts General Hospital in Boston, Massachusetts. Each patient's sputum was cultured under 21%, 50%, and 100% oxygen conditions using the artificial sputum medium, with 0.5 mL aliquots taken from each culture at 24 h, 48 h, and 72 h of culture time for testing. Cultures were photographed when extractions were made to track visual changes. In addition, a 0.5 mL aliquot of each primary sputum sample was taken prior to culturing. This resulted in 10 discrete samples per patient and a final N of 500 samples. Of these, sputum from 11 patients (11 uncultured sputa, 11 cultured sputa from 21% oxygen at 48 h of incubation) underwent nucleic acid extraction17, sequencing libraries were generated using a commercial DNA library preparation kit, and metagenomic sequencing was performed on a whole genome sequencing platform targeting ~ 5 Gb of sequence per sample with 150 base pair, paired-end reads. Raw reads were processed using the bioBakery suite of tools18, which includes quality control and removal of human "contaminant" sequences and taxonomic profiling with the MetaPhlAn3 profiler19. At the time of nucleic acid extraction, 10 million cells of Imtechella halotolerans, a halotolerant species normally found in estuary ecosystems and not in human microbial communities, were spiked into each sample, allowing the quantification of absolute microbial load for each sample20.
Figure 2 shows individual and average outflow oxygen measurements and pH levels over the course of the culture process for 50 sputum samples cultured under each oxygen condition and an example of a visual differential culture phenotype. Cultures were maintained at 37 °C except during brief periods when sparging and removal of sample aliquots was performed. With both sparging intervals of 12 h and 24 h, elevated oxygen concentrations were maintained, although a drop over time was observed for all three oxygen conditions, with 100% oxygen falling to approximately 85%, 50% oxygen falling to 40%, and 21% oxygen falling to 18%. Oxygen conditions remained distinct, and importantly, elevated oxygen concentrations were maintained throughout the process for hyperoxic samples. pH measurements showed a greater degree of variability but stayed well within a physiologically normal range with no statistically significant changes over time. These measurements indicate that these methods maintain discrete differential oxygen conditions throughout the culture process. Lastly, an example of one of many visual culture phenotypes that differentiated across oxygen concentration is shown. This sample had marked turbidity differences after 72 h of culture, with higher oxygen associated with lower visual turbidity. Differential culture phenotypes support the presence of hyperoxia-induced effects on culture communities.
Figure 3 compares microbial load, microbial diversity, and microbial community composition between uncultured sputum and cultured sputum (21% oxygen condition for a period of 48 h). Measurements reveal the only major difference introduced by the act of culturing to be an approximately 20-fold increase in microbial load as compared to uncultured sputum. The immune system and the typical mechanical sputum clearance mechanisms such as coughing normally serve as a regulatory process limiting the microbial load in the lung, even in cases of dysfunction and infection like those seen in pwCF. Ex vivo culture has no such regulatory mechanisms, and microbial communities are instead free to proceed toward cellular saturation. Alpha and beta diversity metrics indicate that despite this difference in microbial load, the underlying community composition remains well-preserved, with minimal global differences introduced by the culture process.
Figure 4 expands on the comparison between uncultured and cultured sputum samples, looking at the binary presence/absence of the 120 microbial species conclusively identified by shotgun metagenomics sequencing from cultured and uncultured sputum obtained from 11 patients. Microbes are clustered based on phylogenetic similarities. 46 (38.3%) of these species were identified in both uncultured and cultured samples (cyan color), while 35 (29.2%) were exclusively identified in uncultured samples (yellow) and 39 (32.5%) were exclusively identified in cultured samples (blue). It is likely that there is greater parity than what we identified using sequencing in terms of what is present and what is absent, but some taxa fall below the sequencing detection threshold in some cases. The differences do indicate that the culture process introduces some bias in cultured compared to uncultured sputum. Most notably, culturing increases the presence of fungi such as Candida and Aspergillus, as well as Enterobacterales members including Escherichia, Serratia, and Streptococcus members. Conversely, Bacteroidetes members such as Prevotella and Clostridiales, which are anaerobes, were present in uncultured samples but not present in cultured samples. This may be attributed to the lack of an anaerobic condition in our experimental model.
Figure 5 shows absorbance-based growth curves of common CF lung pathogens isolated from sputum obtained from 50 different pwCF. These isolates represent phenotypically different clinical isolates obtained using enrichment culture procedures from the Massachusetts General Hospital Clinical Microbiology Laboratory, and include Pseudomonas aeruginosa (N = 53), Staphylococcus aureus (N = 37), Stenotrophomonas maltophilia (N = 12), Klebsiella pneumoniae (N = 3), and Achromobacter sp (N = 7). Growth curves were obtained by culturing each isolate in artificial sputum media at 37 °C in the dark, with ASM sans bacterial inoculation serving as a negative control. The transparent quality of ASM (which is due to filter rather than heat sterilization) allows for conducting optical measures to estimate growth curves. Optical readings at 600 nm (OD600) were taken every 10 min, and the first 24 h of each curve are shown. The absence of changes in optical readings in the ASM-only negative control indicates cultures free of contamination. The demonstrative curves shown here follow typical growth curve patterns indicating the viability of this ASM recipe as a medium for the absorbance-based generation of growth curves.
| Column 1 | Column 2 | Column 3 | Column 4 | Column 5 | Column 6 | Column 7 | Column 8 | Column 9 | |
| Value | Comstock | Kirchner | Sriramulu | Palmer | Flynn | Gallagher | Lai | Source | |
| Mucin | 2% w/v | 0.5% w/v | 0.5% w/v | – | 1% w/v | 2% w/v | 1% w/v | Flynn | |
| Sodium Chloride | 85.5 mM | 85.5 mM | 85.5 mM | 66.6 mM | 89.8 mM | 85.5 mM | 152.3 mM | Lapierre | |
| Potassium Chloride | 29.5 mM | 29.5 mM | 29.5 mM | 15.8 mM | – | 2.95 mM | 15.8 mM | Palmer | |
| Magnesium Sulfate | – | – | – | 0.6 mM | 1 mM | 1 mM | 0.61 mM | Palmer | |
| Iron Sulfate | – | – | – | 0.0036 mM | – | – | – | – | |
| Ammonium Chloride | – | – | – | 2.3 mM | 60 mM | – | – | – | |
| Monopotassium Phosphate | – | – | – | 2.5 mM | 60 mM | – | – | – | |
| Glucose | – | – | – | 3.2 mM | 13 mM | 40 mM | 0.7 mM | Sambeek | |
| Lactate | – | – | – | 9 mM | – | – | – | – | |
| Essential Amino Acids | 14.45x | 0.25 g/L | 0.25 g/L | Per Acid | 0.5x | 0.375x | 1.29x | Palmer | |
| Non-Essential Amino Acids | 28.9x | 0.25 g/L | 0.25 g/L | Per Acid | 0.25x | 0.5x | 8.01x | Palmer | |
| Vitamins | – | – | – | – | – | 1x | 1x | Gallagher | |
| Trace Metals | – | – | – | – | 1x | 1x | 1x | Flynn | |
| Egg Yolk | 0.25% | 0.25% | 0.25% | – | – | 0.25% | 0.25% | Kirchner | |
| Ferritin | 0.0003 g/L | – | – | – | – | 0.0004 g/L | 0.0004 g/L | Gallagher | |
| Salmon Sperm DNA | 1.4 g/L | 4 g/L | 4 g/L | – | – | 1.4 g/L | – | – | |
| DPTA | – | – | 0.0059 g/L | – | – | – | – | – | |
| pH | – | 6.9 | – | 6.8 | – | – | 6.3 | Lapierre | |
| Storage | 0 | 4° | – | – | 4° | 4° | 4° | Kirchner | |
| Sterilization | Autoclave | Filter | Autoclave | – | Autoclave | Autoclave | Filter | Kirchner | |
Table 1: Artificial sputum medium recipe derived from review of literature. (Column 1) Reagents and key values in the formulation of artificial sputum medium. (Columns 2–7) Comparison of recipes from extant literature8,9,10,12,14,15. (Columns 8–9) Artificial sputum medium recipe detailed in this protocol and the corresponding sources that informed each selected value10,11,12,13,14,15.
| Column 1 | Column 2 | Column 3 |
| CF Sputum | ASM | |
| Total Amino Acids | 10.25 mM | 10.76 mM |
| Alanine | 0.96 mM | 0.80 mM |
| Arginine | 0.17 mM | 0.94 mM |
| Asparagine | 0.91 mM | |
| Aspartic Acid | 0.45 mM | 0.80 mM |
| Cysteine | 0.09 mM | 0.33 mM |
| Glutamic Acid | 0.84 mM | 0.80 mM |
| Glycine | 0.65 mM | 0.80 mM |
| Histidine | 0.28 mM | 0.35 mM |
| Isoleucine | 0.60 mM | 0.52 mM |
| Leucine | 0.87 mM | 0.52 mM |
| Lysine | 1.15 mM | 0.64 mM |
| Methionine | 0.34 mM | 0.13 mM |
| Ornithine | 0.36 mM | |
| Phenylalanine | 0.29 mM | 0.26 mM |
| Proline | 0.90 mM | 0.80 mM |
| Serine | 0.78 mM | 0.80 mM |
| Threonine | 0.58 mM | 0.52 mM |
| Tryptophan | 0.07 mM | 0.06 mM |
| Tyrosine | 0.43 mM | 0.26 mM |
| Valine | 0.60 mM | 0.52 mM |
Table 2: Amino acid concentrations previously described in cystic fibrosis sputum and in artificial sputum medium recipe detailed in this protocol. (Column 1) Key amino acids. (Column 2) Amino acid concentrations of sputum from people with cystic fibrosis12. (Column 3) Amino acid concentrations in artificial sputum medium detailed in this protocol
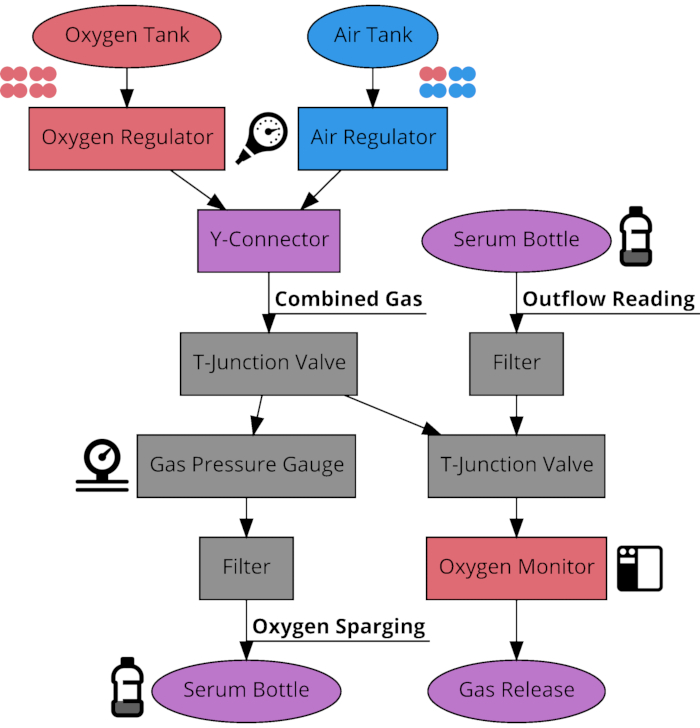
Figure 1: Connection schematic of oxygen sparging system components. Flow diagram of the connections between components of the system used to sparge serum bottles to desired oxygen concentrations between 21% and 100%. The system has 3 modes of use determined by the position of the two T-junction valves. The system can route gas from the tanks through the gas output or through the oxygen percentage monitor, as well as route outflow gas from previously sparged serum bottles through the monitor to check concentration after time has elapsed. Please click here to view a larger version of this figure.
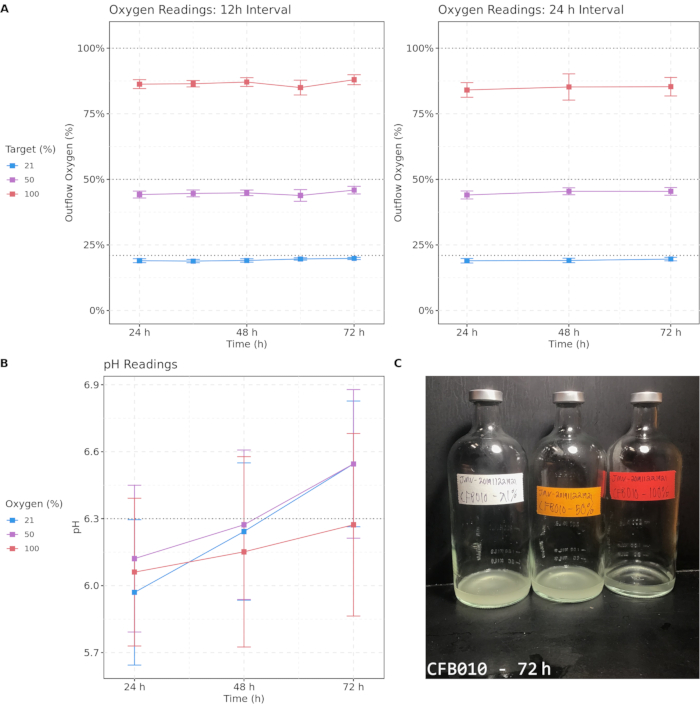
Figure 2: Target oxygen concentrations are approximately maintained with both 12 h and 24 h sparging intervals, and pH remains in the physiological range during culture. (A) Outflow oxygen readings from 12 h and 24 h oxygen sparging intervals over a 72-h period. (B) pH readings for samples measured every 24 h. (C) An example image of cultured sample CFB010 after 72 h, displaying differential turbidity across oxygen concentrations. The color indicates target oxygen percentage; error bars denote 95% confidence intervals. Critical thresholds are emphasized with dashed lines. Please click here to view a larger version of this figure.
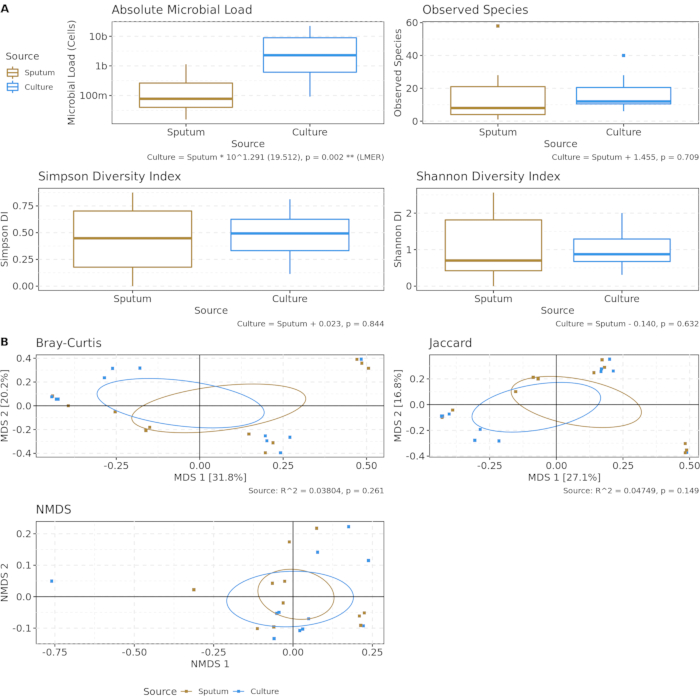
Figure 3: Culturing increases microbial load, but underlying community composition is preserved. Uncultured sputum (yellow) and cultured sputum (blue) using artificial sputum medium at 21% oxygen for 48 h. Aliquots underwent nucleic acid extraction and shotgun metagenomics sequencing to detect possible bias introduced from culture conditions. (A) Absolute microbial load (determined by spike-in controls) and alpha diversity metrics. Using linear mixed effects models, uncultured vs. cultured sputum predicted microbial load but not alpha diversity. (B) Ordination of first two components of beta diversity metrics, controlling for difference in microbial load. No significant difference in either metric after PERMANOVA. Please click here to view a larger version of this figure.
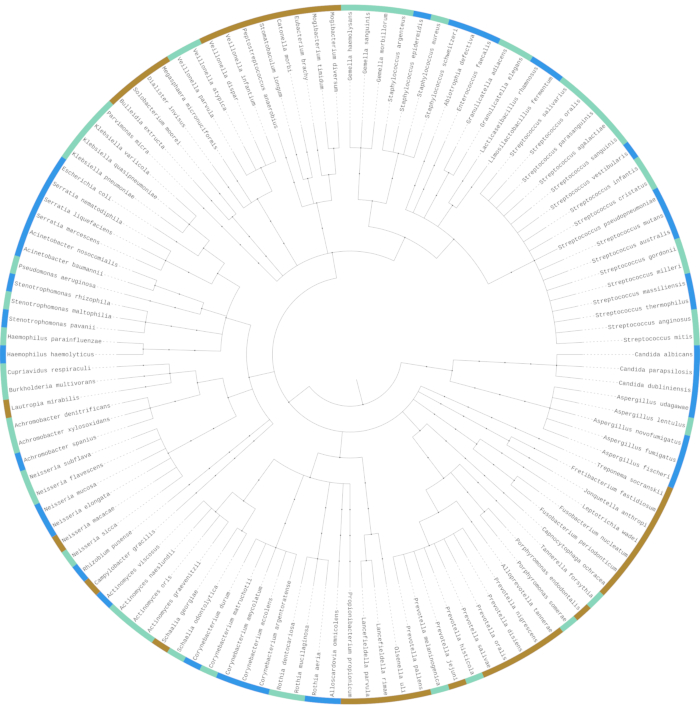
Figure 4: The majority of identified taxa are present in both source sputum and culture, while others only appear in source sputum or culture. Shotgun metagenomics sequencing used to compare differences in microbial community composition between uncultured and cultured sputum samples. Phylogenetic tree of all identified microbial species in sequenced samples (N = 120). Species marked with yellow (N = 35, 29.2%) were only seen in uncultured sputum samples. Species marked with blue (N = 39, 32.5%) were only seen in artificial sputum medium culture samples. Species marked with cyan (N = 46, 38.3%) were seen in both uncultured and cultured samples. Please click here to view a larger version of this figure.
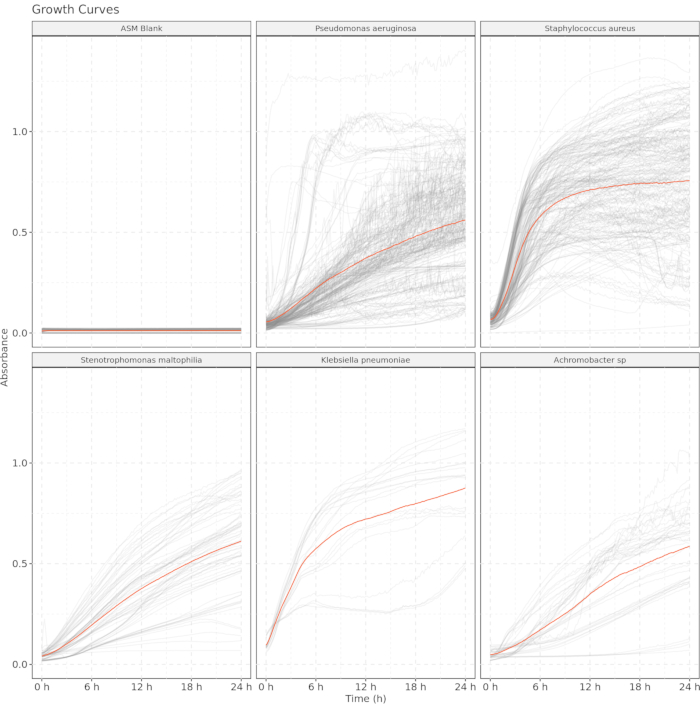
Figure 5: Artificial sputum medium is sufficiently transparent to be used as a growth curve medium for culturing clinical isolates. Optimal density readings at 600 nm were taken every 10 min, and the first 24 h of each curve are shown. Gray lines represent individual readings, and orange lines represent the mean absorbance for each taxon. Artificial sputum medium blank included as a control. Please click here to view a larger version of this figure.