Fission yeast cells expressing the ring marker, Rlc1-GFP (green, Figure 2) and spindle pole body marker Sad1-mCherry (red, Figure 2) were imaged during cytokinesis. Onset of spindle pole body marker (white arrow, Figures 2A, B) is considered as time 0. Rlc1-GFP signal appears at time -4 min with reference to spindle pole body separation (red arrow, Figures 2A, 2B). Rlc1-GFP signal forms a continuous ring 10 min post spindle pole body separation (open arrow, Figures 2A, B) marking the end of actomyosin ring assembly. The actomyosin ring (Rlc1-GFP) starts to decrease in width 22 min after completion of ring assembly and 32 min after spindle pole body separation (closed arrowhead, Figures 2A, B). Ring constriction ends 20 min after onset of constriction, 42 min since onset of ring assembly and 52 min since spindle pole body separation (white asterisk, Figures 2A, B). Finally, cell abscission occurs 72 min post spindle pole body separation (red arrowhead, Figures 2A, B).
The localization of the ring marker Rlc1-Tomato and septum membrane marker Bgs1-GFP were analyzed at the division site throughout cytokinesis. 3D reconstruction of the ring revealed that actomyosin ring marked with Rlc1-Tomato assembled before Bgs1-GFP recruitment (Figure 3, 10 min). During the ring maturation phase Bgs1-GFP was recruited to the division site and overlapped with the actomyosin ring as shown by Rlc1-Tomato (Figure 3, 30 min). This indicates that Bgs1 localizes to the ring upon recruitment. As the ring constricts, Bgs1-GFP also localizes to the ingressing membrane adjacent to the constricting ring (Figure 3, 40 min). At the end of constriction, the Bgs1-GFP signal is visible at the ingressed membrane barrier (Figure 3, 50 min). Finally, after constriction the Rlc1-Tomato signal is absent, while Bgs1-GFP appears to localize throughout the membrane barrier with a disc like appearance (Figure 3, 60 min). Thus these observations indicate that the septum synthesizing enzyme Bgs1 localizes to the ring after assembly and persists after ring constriction at the membrane barrier as has been reported before17.
The distribution of proteins along the actomyosin ring was analyzed to determine efficiency of cytokinetic events (Figure 4). Uneven or nonhomogenous distribution of proteins along the actomyosin ring has been associated with impaired signaling and inefficient cytokinetic ring assembly26. Here we show two 3D reconstructed rings during the maturation phase, expressing the F-Bar protein Cdc15-GFP (Figure 4). The image on the left shows even distribution of Cdc15-GFP along the ring with a low coefficient of variance (0.098), while the image in the right shows uneven distribution with a high coefficient of variance (0.226, Figure 4).
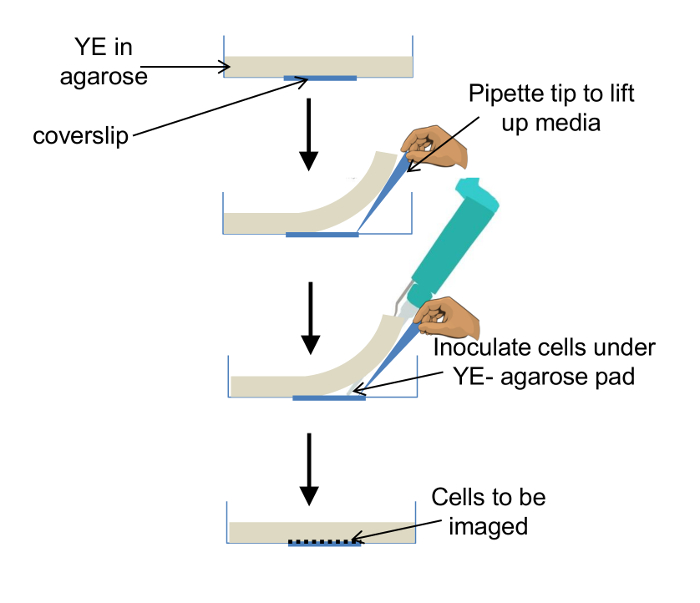
Figure 1: Preparation of Culture Dish for Imaging. Schematic describing inoculation of cells into a glass bottomed culture dish overlaid with media pad. See protocol for details (Section 1). Please click here to view a larger version of this figure.
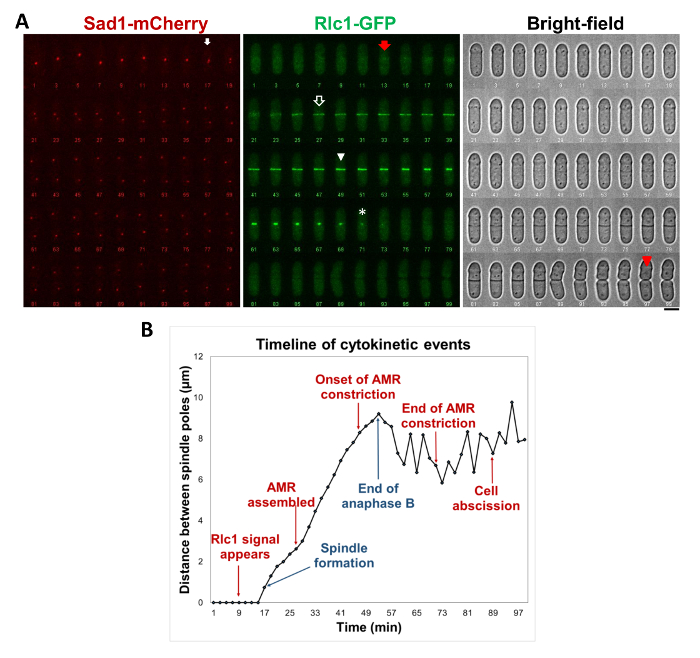
Figure 2 Timing of Cytokinetic Events in S. pombe. (A) Montage of a cell expressing spindle pole body marker Sad1-mCherry (left panel), ring protein Rlc1-GFP (middle panel) and bright-field (right panel) undergoing cytokinesis is shown at 2 min intervals. White arrows marks spindle pole body (centrosome) separation at 17 min, red arrow marks initial recruitment of Rlc1-GFP at the division site, open arrow marks ring maturation at 27 min, arrow head marks onset of ring constriction at 47 min, asterisks marks end of ring constriction at 69 min and red arrowhead marks cell abscission at 97 min. (B) Timeline of cytokinetic events with reference to mitosis as determine by spindle pole body formation and separation. Cells were imaged at 25 °C. Please click here to view a larger version of this figure.
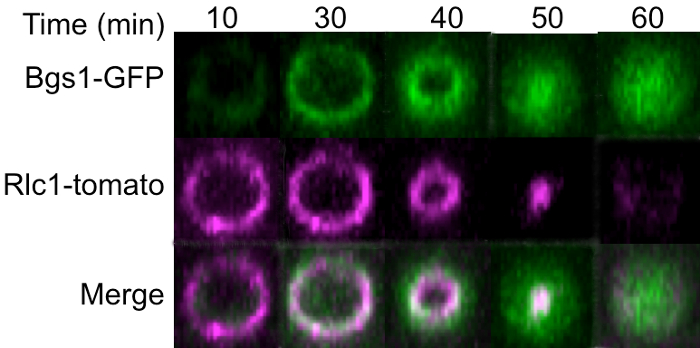
Figure 3: Cell Division Site During Different Stages of Cytokinesis. 3D reconstruction of actomyosin ring Z-stacks during different stages of cytokinesis for the same cell, ring assembly (10 min post Spindle pole body separation), ring maturation (30 min post Spindle pole body separation), ring constriction (40 min post Spindle pole body separation), end of constriction (50 min post Spindle pole body separation), septum barrier (60 min post Spindle pole body separation). Rlc1-Tomato marks ring while Bgs1-GFP marks the plasma membrane enveloping the septum. Scale bar = 5 µm. Please click here to view a larger version of this figure.
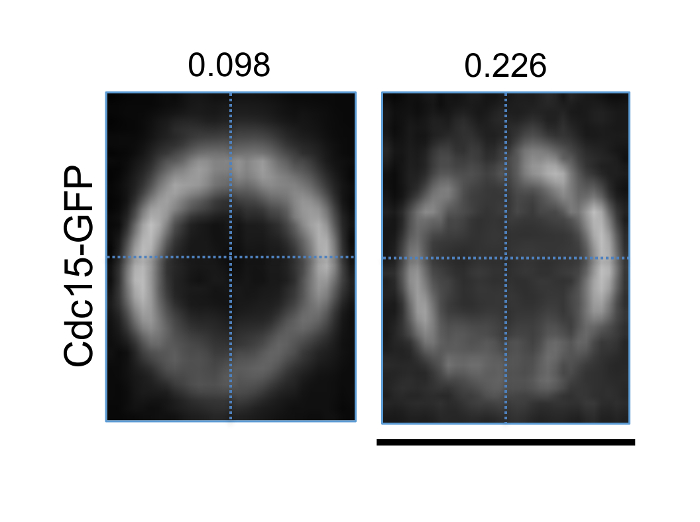
Figure 4: Distribution of Proteins along the Ring. Images of 3D reconstructed actomyosin ring from wild type cells expressing Cdc15-GFP divided into four quadrants. The co-efficient of variance of each ring is stated. Scale bar = 5 µm. Please click here to view a larger version of this figure.
