The devised protocol enabled MALDI-IMS analysis of E. precatoria and E. edulis seeds. As a result, we could confirm carbohydrates' molecular weight and degree of polymerization (DP) as a partial structural elucidation. The molecular information provided by the MALDI-IMS analysis (Figure 1 and Figure 2) exhibited peaks representing [M+K]+ adducts of hexose oligomers (Δ = 162 Da) without adding salt to the matrix. Hexose dimers (m/z 381), trimers (m/z 543), tetramers (m/z 705), pentamers (m/z 867), hexamers (m/z 1029), and up to 14-unit oligomers (m/z 2325) were identified in both seed tissues. The box plots for both samples (Figure 3) indicate the peak intensity of each hexose oligomer found in the seed endosperm, demonstrating their distributions and a slightly higher content of high DP oligomers.
Previously, we provided EDS data on E. oleracea seeds, which indicated that potassium adducts detected in the MALDI-IMS analysis were due to the sample's intrinsic characteristic of elevated potassium content10. This study also analyzed E. precatoria and E. edulis seeds to identify the main elements in the sample's sections (Supplementary Figure S2 and Supplementary Figure S3). The atomic composition observed on the tissue surface was homogeneously distributed, showing mainly nonmineral elements, such as carbon and oxygen, and lower mineral content. In both samples, potassium was the main mineral element found by EDS analysis (Supplementary Figure S2 and Supplementary Figure S3).
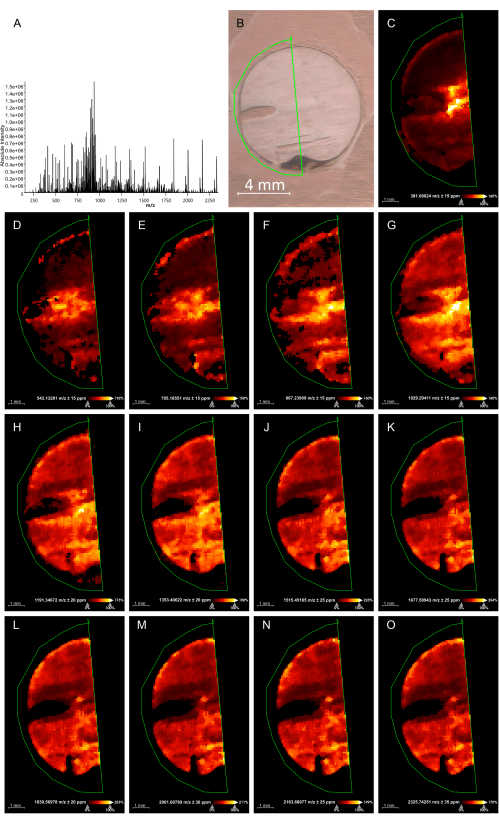
Figure 1: Hexose oligomer distribution in Euterpe precatoria seeds by MALDI-IMS in positive mode. (A) Mass spectrum obtained. (B) Histological image and the frame used for the MALDI-IMS analysis. (C–O) Image representations of a specific m/z signal describing its intensity in the tissue for each [M+K]+ adduct from hexose dimer (C), trimer (D), up to 14 units (E–O) in the seeds' endosperm. Scale bar = 4 mm (B). Abbreviation: MALDI-IMS = matrix-assisted laser desorption/ionization-imaging mass spectrometry. Please click here to view a larger version of this figure.
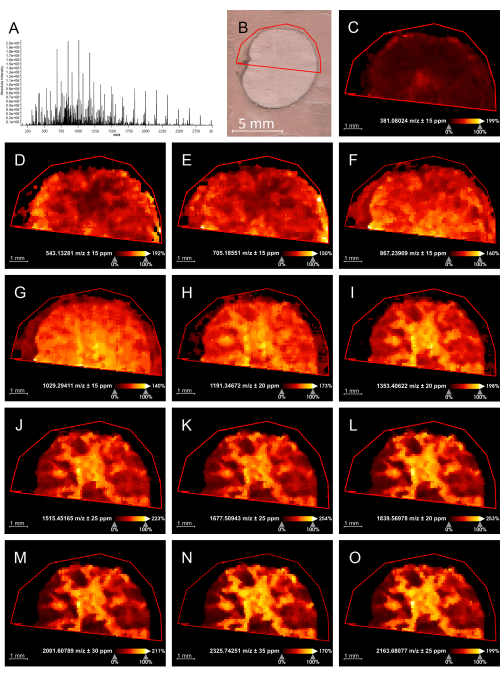
Figure 2: Hexose oligomer distribution in Euterpe edulis seeds by MALDI-IMS in positive mode. (A) Mass spectrum obtained. (B) Histological image and the frame used for the MALDI-IMS analysis. (C–O) Image representations of a specific m/z signal describing its intensity in the tissue for each [M+K]+ adduct from hexose dimer (C), trimer (D), up to 14 units (E–O) in the seeds' endosperm. Scale bar = 5 mm (B). Abbreviation: MALDI-IMS = matrix-assisted laser desorption/ionization-imaging mass spectrometry. Please click here to view a larger version of this figure.
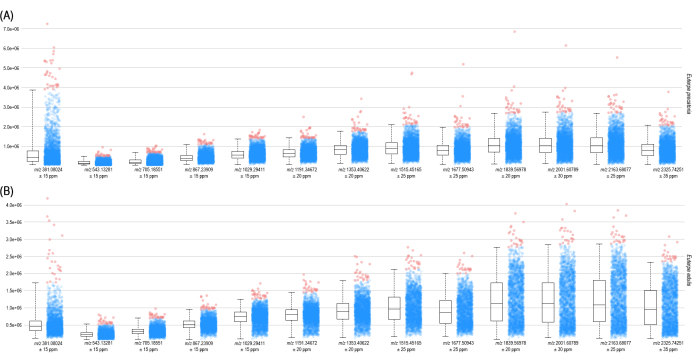
Figure 3: Box plot visualizations. (A) E. precatoria and (B) E. edulis, indicating the peak intensity and distributions of each hexose oligomer found in the seed tissue. Please click here to view a larger version of this figure.
Supplementary Figure S1: Matrix coating template. The DHB matrix was sprayed onto the platform using drawing software for equal distribution on a 75 mm x 25 mm slide of horizontal parallel lines spaced by 1 mm. Please click here to download this File.
Supplementary Figure S2: Elementary composition analysis of E. precatoria seeds by energy dispersive X-ray spectrometry. (A) E. precatoria seed sample surface section. (B) Major elements semi-quantified. The main components correspond to about 47.0% carbon (C), 52.7% oxygen (O), and 0.2% potassium (K). Please click here to download this File.
Supplementary Figure S3: Elementary composition analysis of E. edulis seeds by energy dispersive X-ray spectrometry. (A) E. edulis sample surface section. (B) Major elements semi-quantified. The main components correspond to about 47.7% carbon (C), 53.8% oxygen (O), 0.3% potassium (K), and some traces of chlorine (Cl). Please click here to download this File.
Supplementary Figure S4: Delimitation of the area in an image and data analysis. (A) Delimitation of the area to be analyzed and editing the parameters and raster width to 100 µm. (B) Identification of a matrix cluster and creation of a mass list. (C) Calibration of data and saving the 2D data tool. Please click here to download this File.
