The data is presented as the mean with ± standard error and was obtained from three independent experiments, with at least triplicates in each group. Experimental data, including colony counts, OD600 measurements, and TTC staining results, were graphed and statistically analyzed using graphing and statistical software (see Table of Materials). One-way ANOVA or t-test was used to analyze the data, and a p-value <0.05 was considered significant. Scanning was performed with a 48-bit full-color optical scanner at a resolution of 1200 dpi, and subsequent measurements of the colony diameters were conducted using ImageJ software.
As depicted in the growth curves in Figure 2, petite mutants of C. glabrata C2-1000907 (T3p) exhibited slower growth compared to the untreated parental fungi of regular size (T0). When C. glabrata was mixed with 0.2% RB for 15 min and exposed to 4.38 J/cm2 green light (PDT treatment), the titer of the fungi decreased from 107.5 CFU/mL to 104.5 CFU/mL (at least 3 logs, Figure 3) compared with the control group (RB only), which had a cell density of around 107 CFU/mL. Additionally, after aPDTs, petite colonies were observed. These petite colonies had an average size of 0.4 mm ± 0.25 mm instead of the usual size of 1.5 mm ± 0.8 mm (Figure 4A). Small-size colonies with mitochondrial dysfunction can be identified by their size and color after staining with 20% TTC. The petite colonies that maintained a white color (Figure 3B, white arrows) exhibited mitochondrial dysfunction, while some smaller red-colored colonies and normal-sized colonies retained normal mitochondrial function (Figure 4B). The traditional DNA intercalating agent ethidium bromide (Figure 4C) and fluconazole (Figure 4D) can effectively induce petite mutants of Candida. Disk diffusion is a simple method to define drug susceptibility in fungi. In Figure 4D, a clear circular zone of no growth surrounded the fluconazole disk containing 25 µg fluconazole in the T0 culture (without PDT), indicating less drug resistance in this parental fungus. Nonetheless, the C. glabrata C2-1000907 is still defined as a resistant strain by minimal inhibition concentration toward fluconazole (data not shown) and the diameter of the clear zone. In the T3 culture, many petite colonies formed near the disk (arrow, lower panel), suggesting drug resistance after three repeated PDT treatments.
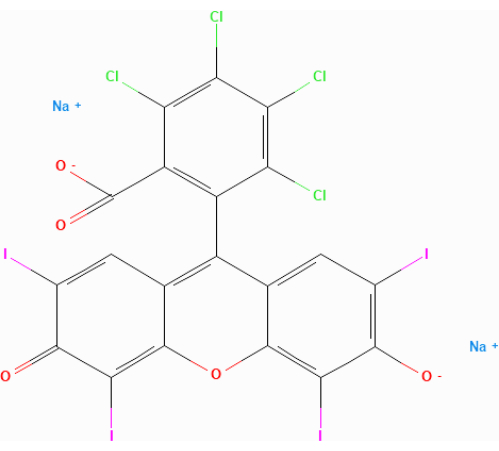
Figure 1: Molecular structure of rose Bengal (RB). Please click here to view a larger version of this figure.
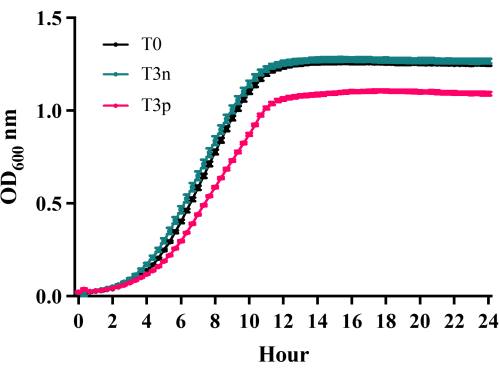
Figure 2: Growth Curves of C. glabrata C2-1000907 24 h following repetitive RB-PDT. Growth curves were compared for three strains: C. glabrata C2-1000907 T0 (naïve parental cells), T3n (colonies with normal size after 3 repeated treatments), and T3p (petite colonies after 3 repeated treatments). Compared to T0 and T3n, T3p colonies exhibited a significantly slower growth rate (n = 3, p < 0.05). All cells are grown in YPD medium. Please click here to view a larger version of this figure.
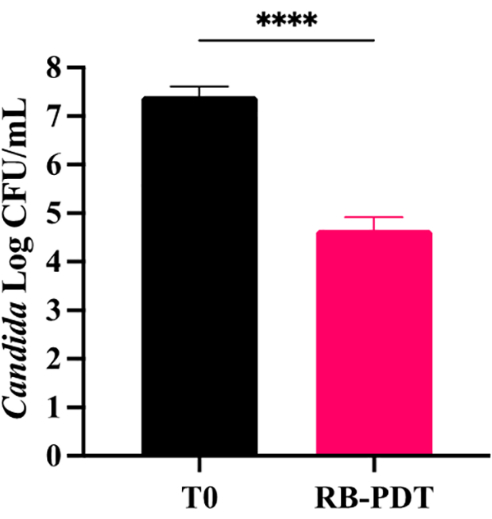
Figure 3: Growth Inhibition of C. glabrata C2-1000907 following aPDT. A single aPDT treatment with a 4.38 J/cm² light dose and 0.2% RB significantly inhibited C. glabrata C2-1000907 growth by 3 logs compared to the naïve control (p < 0.0001, unpaired t-test). Error bars represent mean ± SEM, data is pooled from 3 independent experiments. Please click here to view a larger version of this figure.

Figure 4: Mitochondrial dysfunction analysis using triphenyltetrazolium chloride (TCC) staining. (A) C. glabrata C2-1000907, 24 h following aPDT treatment (0.2% RB, 4.38 J/cm² green light), displayed a bimodal colony size distribution, with large (1.5 mm ± 0.8 mm) and small (petite) colonies (0.4 mm ± 0.25 mm). (B) TCC staining revealed that the large colonies were functional, as they turned red, while the petite colonies were non-functional, as they remained white. (C) Petite colonies were also induced by ethidium bromide (100 µg/mL for 30-40 min) and (D) fluconazole (25 µg/mL) treatment. A clear circular zone of no growth surrounded the fluconazole disk containing 25 µg fluconazole in the T0 culture (without PDT), indicating susceptibility. In the T3 culture, many petite colonies formed near the disk (arrow, lower panel), suggesting drug resistance after three repeated PDT treatments. Scale bars: 1 mm. Please click here to view a larger version of this figure.
