1. Preparation of cells
- Target cell culture: Culture of the mouse beta cell lines, NIT-1 and NIT-4, was a previously described1,2 in DMEM supplemented with 10% FBS, 0.02%BSA, Non-essential Amino Acid, 15mM HEPES, 16.5mM Glocose and Penicillin/Streptomycin. For 51Cr labeling, seed cells in a flat-bottom 96-well culture plate at 5X104 cells/well in 200 μl culture medium. For microscopy experiments, seed cells in a 8-well Lab-Tak II chambered coverglass at 5×104 per chamber in 500 μl culture medium, and allow to grow for 48 hours prior to using in the cytotoxicity experiments.
- Control T cell line culture: Use the Jurkat cell line (ATCC, CD3+ human lymphocyte cell line) as a negative control. Culture Jurkat cells in RPMI1640 supplemented with 10% FBS, 2 mM L-glutamine, 1.5 g/L sodium bicarbonate, 10 mM HEPES and 1.0 mM sodium pyruvate.
2. Collection and activation of effector autoreactive CD8+ T cells
NOD.Cg-Rag1tm1MomTg(TcraAI4)1Dvs/DvsJ [NOD-AI4a] and NOD.Cg-Rag1tm1MomTg(TcrbAI4)1Dvs/DvsJ [NOD-AI4b] were purchased from The Jackson Laboratory (Bar Harbor, ME) and bred in our mouse facility. F1 hybrid progeny from matings of NOD-AI4a with NOD-AI4b [NOD-AI4a/b] develop diabetes at 3-5 weeks of age. All mice were housed in specific pathogen-free facilities and approved by the relevant institution’s Animal Care and Use Committee.
- Euthanize 3-4 week old NOD-AI4a/b mice using CO2.
- Collect spleens from mice. Euthanized mice are prepared with ethanol spray and opened under surgical hood. Spleens are removed and put in ice-cold HBSS buffer immediately.
- Collect spleen cells using a 7 mL glass homogenizer. Two to three spleens each are put in 5 mL of cold HBSS and homogenized with a 7 mL glass homogenizer. Homogenate is collected and centrifuged at 1200rpm for 5 minutes at 4°C using a Sorvall RT7+ centrifuge. Cell pellets are collected.
- Remove red blood cells using hypotonic solution treatment. Pellets are treated with 5 mL hypotonic solution/spleen on ice for 5 minutes, then neutralized by adding an equal volume of HBSS. Collect cells by centrifugation, as above, and resuspend in 50 mL HBSS. Pass the suspended cells through cell strainer and count using a hemocytometer with TrypanBlue staining.
- Activate cells. Resuspend cells at 5×106/mL and activate using a 3 day culture with 0.1 μM AI4 mimotope (amino acid sequence YFIENYLEL) and 25 U/mL IL-2 in RPMI1640 supplemented with 10% FBS, 2 mM L-glutamine, 1.5 g/L sodium bicarbonate, 10 mM HEPES and 1.0 mM sodium pyruvate.
3. 51Cr release assay to examine CML
- Label target cells: Add 51Cr to target cells at 1 μCi/well and culture for 3 hours, and wash 3 times with culture medium.
- Suspend effector cells in RPMI 1640 medium as mentioned in step 2.5 so that the final volume added to each well is 200 μl.
- After the final wash of labeled target cells, add activated effector cells to the target cell cultures at desired effector to target (E:T) ratios. We usually use E:T ratios at 0.4:1, 1:1, 2:1, 5:1, 10:1, 20:1 and 50:1.
- Add fresh modified RPMI 1640 medium (see step 2.5) to 6 wells of target cells to act as a spontaneous lysis control.
- Co-culture effector and target cells for 16 hours.
- Collect supernatant in 6×50 mm Lime Glass tubes
- Lyse cells by adding 100μl 2% SDS and pipetting several times gently.
- Collect cell lysate in another set of 6×50 mm Lime Glass tubes.
- Wash wells once with clean H2O and add the wash to the cell lysate tubes.
- Count the supernatants and cell lysates using a gamma counter.
- Calculate specific lysis as follows:
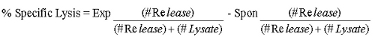
4. Staining of cells for live cell imaging
Among most commonly used mitochondrial membrane potential dyes, TMRM exhibits the least mitochondrial toxicity.3 Therefore, we choose TMRM for these live cell imaging studies.
- Prepare a 40 mM stock TMRM solution in DMSO and store at -20°C. Make a [25 nM] fresh working solution in phenol red-free DMEM with all the supplements for NIT-1 cell culture 1.
- Stain target NIT-1 cells with TMRM 25 nM for 30 min at 37°C.
- Replace the media with fresh media containing 5 nM TMRM to maintain the equilibrium distribution of the dye4.
- Count activated effector AI4 T cells using a hematocytometer.
- Stain activated effector AI4 T cells with 1 μl/mL Picogreen for 5 minutes at room temperature and wash with phenol red-free culture medium 3 times.
5. Assessing T lymphocyte-mediated beta cell death using live cell imaging
- Mount the chambered coverglass with stained NIT cells on the stage of a Zeiss LSM 510 confocal microscope. Remove the tab with a dremmel and gas sterilize the chambered glass with etheline oxide. The stage is equipped with a live-cell imaging chamber that has temperature controlled at 37°C and 5% CO2 with moisture.
- Add activated and stained T cells to designated chambers at different E:T ratios.
- Added same number of stained Jurkat cells to the control chambers.
6. Obtaining live cell images
The microscope is equipped with a motorized stage that allowed us to repeatedly acquire images at different locations from the same or different chambers automatically over the time course of the experiment.
- Using a 63x oil objective lens, take images at an interval of every 3 minutes for a total of 300 frames per location.
- Detect TMRM staining using an excitation wave length of 543 nm and filter set for emission 565-619 nm
- Detect Picogreen staining using an excitation 488 nm and a filter set for emission 500-630 nm. The video was created using Zeiss LSM software.
7. Video conversion for publication
- Using the Zeiss LSM software, create an uncompressed movie file in an AVI format.
- To convert the video into the published format, import the video into the software package FIJI (http://pacific.mpi-cbg.de/wiki/index.php/Fiji), an ImageJ (http://rsb.info.nih.gov/ij/) distribution, using the Bioformats plugin from LOCI (http://www.loci.wisc.edu/software/bio-formats).
- Adjust the brightness and contrast settings to balance the green and red signals and the frame rate was set to 2 frames/second using the animation options menu.
- Crop the video to 512 x 512 pixels and export as an uncompressed AVI file.
- Open this AVI file in the software Quicktime 7 (Apple computer, Cupertino, CA) and compress and export in the open video file format H264/MPEG-4 at 1200 kbs/second.
8. Representative Results:
- A representative result of 51Cr CML assay. Figure 1 shows a representative result of a CML assay where percent specific lysis (Y axis) increases as the Effector:Target ratio (X axis) increases. Target cells are primary islets isolated from immunodeficient NOD.Rag1-/- mice and labeled with 51Cr. Splenocytes from NOD.AI4α/β transgenic mice are isolated and activated as described in step 2 above. Targets and effectors were co-cultured for 16 hours. Percent specific lysis is calculated as described in step 3.11 above.
- Two representative (positive and negative) results of a T cell-mediated beta cell mitochondrial membrane potential dissipation are presented. Cells are stained and reactions are recorded as described in steps 4, 5 and 6 above, video files are created as in step 7. Video 1: The mouse beta cell line NIT-1 was stained the mitochondrial membrane potential dye TMRM (Red). Activated, autoreactive CD8+ T cell AI4 (Stained green with Picogreen) were co-cultured with these NIT-1 cells at an E:T ratio of 50:1. NIT-1 mitochondrial membrane potential dissipated gradually during a 400-minute duration, but only in the clusters that were interacting with AI4 T cells. Video 2: As a control experiment, NIT-1 cells were stained with TMRM and co-cultured with Picogreen stained Jurkat cells at an E:T ratio of 50:1. Jurkat cells are a human T cell line that is MHC mismatched. NIT-1 cells maintained mitochondrial membrane potential throughout the entire duration of the 400 minutes co-incubation period. Jurkat cells did not interact with NIT-1 clusters and therefore, do not induce killing.
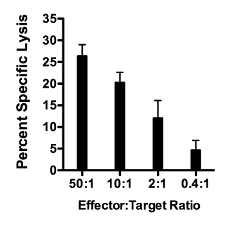
Figure 1. Representative result of CML using as target cells primary islets isolated from immunodeficient NOD.Rag1-/- mice and splenocytes from NOD.AI4α/β transgenic mice. Targets and effectors were co-cultured for 16 hours. Percent specific lysis increases as the Effector:Target ratio increases.
Video 1. Mouse beta cell line NIT-1 was stained the mitochondrial membrane potential dye TMRM (Red). Activated autoreactive CD8+ T cell AI4 (Stained green with Picogreen) were co-cultured with these NIT-1 cells at E:T ratio of 50:1. NIT-1 mitochondrial membrane potential dissipated gradually during a 400-minute duration, but only in the clusters that were interacting with green AI4 T cells. Click here to watch video.
Video 2. NIT-1 cells were stained same as described in Video 1 and co-cultured with Picogreen stained human T cell line Jurkat. NIT-1 cells were able to maintain mitochondrial membrane potential throught out the duration of 400 minutes. Jurkat cells did not interact with NIT-1 clusters and therefore do not induce killing. Click here to watch video.
