A representative example of an insect mortality assay is depicted in Figure 3. In this experiment, insects were injected with approximately 50 colony forming units (CFU) of either wild type (ATCC19061) or an attenuated mutant strain (lrp13) of Xenorhabdus nematophila grown to mid-log phase (n=6 insects per strain). Insects were observed for approximately 72 hr, and the percent of injected insects still alive at each timepoint recorded. In this case, the attenuated strain exhibited a clear delay in insect killing; the wild-type strain killed all 6 larvae within 30 hr post-injection, prior to the death of any mutant-infected larva.
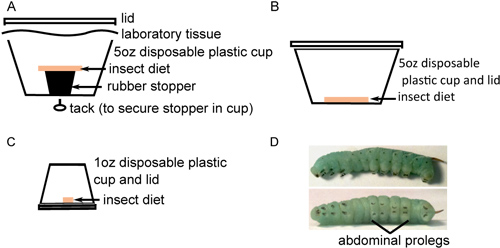
Figure 1. Insect rearing in preparation for injection. A) About 40 surface-sterilized eggs are placed at the bottom of a 5 oz cup with sterile insect diet resting on a rubber stopper. B) Twenty newly hatched insects are transferred to 5 oz cups with sterile insect diet on the bottom and incubated for 2 days. C) Insects are next transferred individually to 1oz cups with sterile diet on the bottom and incubated until they mature. D) Fourth instar M. sexta larvae with prominent stripes along the body (top) and black crochets on the abdominal prolegs (bottom).
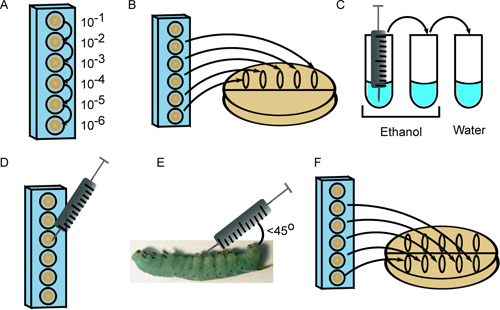
Figure 2. Injection of 4th instar M. sexta larvae. A) Bacteria are serially diluted in a 96-well plate. B) Ten microliters of multiple dilutions are plated to enumerate the inoculum. C) The syringe is sterilized with 3 rinses in ethanol (2x) and sterile water. D) Ten microliters from the appropriate dilution are drawn into the syringe. E) The cell suspension is injected at a 45° angle behind the first abdominal proleg. F) Dilutions are again plated to provide a second measure of the inoculum.
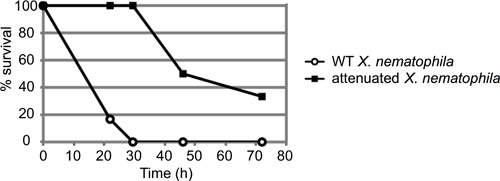
Figure 3. Representative result of M. sexta injection assay. About 50 colony-forming units (CFU) of Xenorhabdus nematophila cells in stationary phase (10 μl from the 10-4 dilution) were injected into six 4th instar M. sexta larvae per strain. Both wild type and a mutant strain (lrp) with an established virulence defect were injected and the insects monitored for mortality over time. Results are reported as percent surviving insects over time (in hours). These curves are statistically distinct, with a p-value of 0.000458 via log-rank analysis.
