Isolation of adult murine cardiomyocytes from genetically engineered mice to study the function of specific genes of interest in vitro has become a powerful tool to further understand cardiac pathophysiology. This method is currently used by only a small but increasing number of basic science laboratories worldwide. However, isolation of adult ventricular murine cardiomyocytes can be tricky and needs to be done thoroughly and repetitively by experienced hands. Figure 1 shows freshly isolated exemplar atrial and ventricular cardiomyocytes. For ventricular cardiomyocytes we recommend to only use rod- shaped, striated ventricular myocytes of healthy appearance with no spontaneous contractions. Compared with ventricular myocytes, atrial myocytes are shorter and thinner. The characteristic striation and rod-shape of ventricular cardiomyocytes is missing in adult atrial murine cardiomyocytes. The yield for ventricular cardiomyocytes isolated from an intact mouse heart is 5×105 – 1×106. For atrial cardiomyocytes it is significantly less. We generally expect to isolate about 5 – 25,000 atrial cells from one mouse heart. Once isolated, cardiomyocytes can be subjected to different in vitro techniques including electrophysiological recordings (Figure 1). We recommend to use isolated cardiomyocytes within the first 6 hr after isolation. Figure 1 shows exemplar Kv channel outward recordings (on the right) of atrial and ventricular cardiomyocytes evoked by different depolarization steps by whole-cell patch clamp technique. The characteristic shape of murine adult ventricular and atrial Kv channel repolarizing currents can be seen over a defined time course (here 4 sec, Figure 1). Please note that the current amplitude is significantly lower in murine adult atrial cardiomyocytes compared to ventricular cardiomycytes.
| Solution | Contents (in mM, if not specified differently) |
| 1 | 117 NaCl, 4 KCl, 10 HEPES, 1 KH2PO4, 4 NaHCO3, 1.7 MgCl2, 10 Glucose |
| 2 | 117 NaCl, 4 KCl, 10 HEPES, 1 KH2PO4, 4 NaHCO3, 1.7 MgCl2, 10 Glucose + 229.5 μM EGTA |
| 3 | 117 NaCl, 4 KCl, 10 HEPES, 1 KH2PO4, 4 NaHCO3, 1.7 MgCl2, 10 Glucose, CaCl2 0.1 + 0.8 g/L Collagenase Type 2 (300 U/L)* |
| 4 | 117 NaCl, 4 KCl, 10 HEPES, 1 KH2PO4, 4 NaHCO3, 1.7 MgCl2, 10 Glucose, CaCl2 0.2 + 0.8 g/L Collagenase Type 2 (300 U/l)* + BSA (1 g/L) |
| 5 | 117 NaCl, 4 KCl, 10 HEPES, 1 KH2PO4, 4 NaHCO3, 1.7 MgCl2, 10 Glucose, CaCl2 0.5 + BSA (1 g/L) |
| 6 | 117 NaCl, 4 KCl, 10 HEPES, 1 KH2PO4, 4 NaHCO3, 1.7 MgCl2, 10 Glucose, CaCl2 1.0 + BSA (1 g/L) |
Table 1. Isolation solutions. Equilibrated for 10 min with Carbogen (95% O2, 5%CO2), at 37 °C pH=7.4 (NaOH). * Activity may depend on batch number, so previous testing of collagenase activity is recommended.
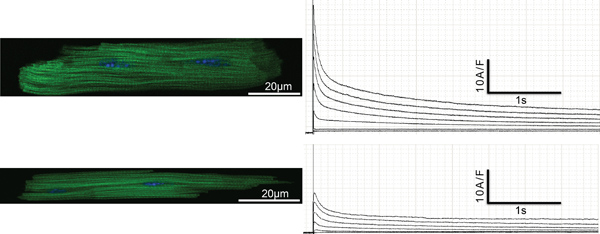
Figure 1. Upper Left: Exemplary ventricular cardiomyocyte stained with DAPI (nuclei, blue) and Alexa Flour 488 Phalloidin (α-actinin, green). Upper Right: Exemplary traces of whole cell patch clamp recording. Lower Left: Exemplary atrial cardiomyocyte stained with DAPI (nuclei, blue) and fluorochrome conjugated Phalloidin (α-actinin, green). Lower right: Exemplary traces of whole cell patch clamp recording.
