Figure 2. The effects of two different HYPR spatial filters on the smoothness of the time-activity data at a single striatal voxel. Top row: 11C-raclopride PET emission images from a 3 minute frame centered at 46.5 minutes (not filtered, filtered with a 3 x 3 x 3 voxel kernel, filtered by a 5 x 5 x 5 voxel kernel). Middle row: 11C-raclopride PET emission images from a 3 minute frame centered at 61.5 minutes (not filtered, filtered with a 3 x 3 x 3 voxel kernel, filtered by a 5 x 5 x 5 voxel kernel). Bottom row: Corresponding time activity curves from the same single voxel location in the left dorsal caudate. Note that the apparent dip in the 11C-raclopride uptake (due to release of dopamine) at the time of smoking is preserved although the noise is diminished with greater filter size.
Figure 3. A selection of representative dopamine response functions that were pre-computed for fitting the lp-ntPET model to the PET time-activity data at each voxel according to Normandin et al. 7. In the case of our smoking paradigm, cigarette smoking commences 45 minutes after tracer injection begins. Even if striatal dopamine responses encode anticipation of smoking – e.g. due to handling of the cigarette or other cues that foretell smoking – we reasoned that the response functions could safely be limited to curves that take off from baseline no earlier than 5 minutes prior to smoking (a). Similarly, curves were limited to take-off times no later than 15 minutes after start of smoking. Curves with take-off times at 40 minutes represent possible dopaminergic responses due to expectation. (b) Representative response functions all taking off from baseline at 45 minutes; the time when smoking commences. 500 different plausible response functions are generated. For illustration, plots in (a) and (b) show only a sampling of curve shapes and take-off times.
Figure 4. (a) The operational equation for the lp-ntPET model. The model is linear in parameters (R1, k2, k2a, γ) which allows fast computation of parameter estimates at each voxel within the striatal mask. (b) Parametric images of (R1, k2, k2a, γ) for a single coronal brain slice for a single subject. Although γ alone is the parameter that encodes the magnitude of a dopamine response, simultaneous estimation of all 4 tracer parameters is necessary to fit the model to the time-activity data at each voxel.
Figure 5. Fits of the conventional (MRTM) and new (lp-ntPET) models to the time-activity data from a voxel in the left caudate. MRTM fit is in blue. lp-ntPET fit is in red.
Figure 6. (a) shows the weighted sum of squared residuals (WSSR) from MRTM and (b) from lp-ntPET fits to the data at each striatal voxel. The two WSSR images produced from the same data are compared to produce a map of the F-ratio at each voxel (i.e. an F-map), shown in (c). (d) The F-map is thresholded at p < 0.05 to produce a binary significance map (see step 2.10 of the protocol). For n timeframes, and 4 parameters of the lp-ntPET model, the threshold for the F-statistic corresponding to a probability level, p < 0.05 (for 90 minutes of data binned in 3-minute frames, the threshold is 4.23) (e) The significance map is filtered with a morphological filter (an “opening”) to eliminate tiny clusters of voxels that are most likely to represent noise. The Final Significance Mask preserves only those voxels in the striatum whose TACs are better fit (statistically) by the lp-ntPET model as opposed to the conventional MRTM model and thus are believed to contain a dopaminergic response to smoking. This threshold does not correct for multiple comparisons. Instead, to guard against false positive findings, we create Final Significance Masks for a control condition as well (see Figure 7 and Protocol Steps 1.8 – 1.10).
Figure 7. (a) shows one coronal slice of the Final Significance Mask for the smoking condition in a single subject. Figure (b) shows the Final Significance Mask for corresponding subject and slice in the baseline condition. The presence of clusters of retained voxels in the mask of smoking as opposed to the near-complete absence of clusters in the mask of control supports the contention that the dopamine movies (see below) are not simply chance events or events related to noise in the data. (Note: the injected activity – and thus the signal to noise ratio – in the baseline and smoking conditions were comparable.)
Figure 8. The dopamine movie of a single slice of brain in the coronal orientation shows the frame-by-frame dopamine level relative to the basal (resting) dopamine level. (a) shows the movie of the baseline condition and (b) shows the movie of the smoking condition. The dopamine levels are encoded in color. Specifically, the colors – shown in the color bar with corresponding numerical values – represent the change in dopamine above the basal level as a percent of basal. Again, the dopamine levels are shown only for voxels in the Final Significance Mask that exceed the p < 0.05 significance level.
Figure 9. The multi-slice, multi-condition dopamine movie for the same subject as in Figure 8 with all slices of the ventral striatum displayed simultaneously for baseline and smoking conditions.
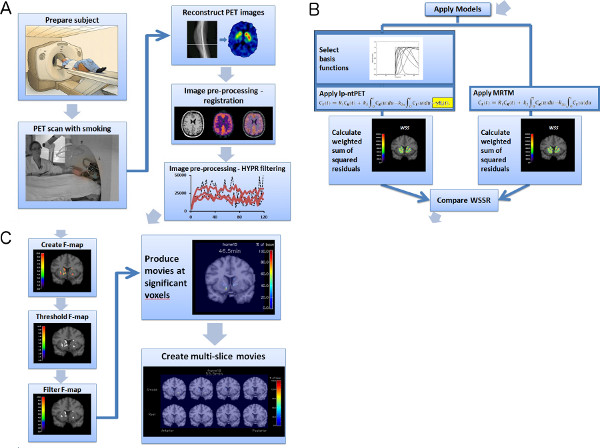
Figure 1. Flowchart of experiment and image analysis procedures (a-c). Click here to view larger figure.
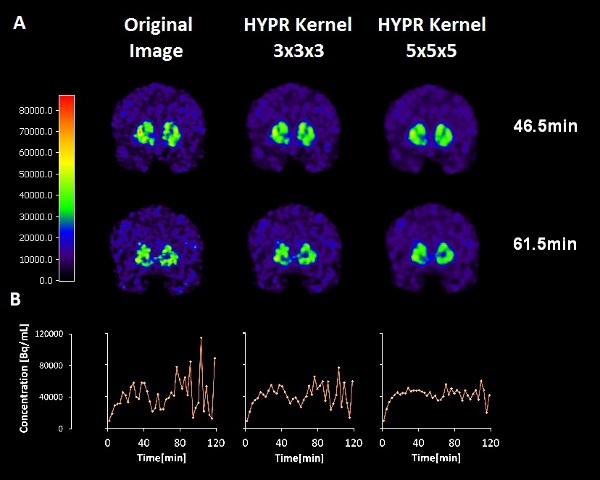
Figure 2. Effects of HYPR filters of different kernel size on images (top and middle) and on time activity curves (bottom) at a single voxel. Click here to view larger figure.
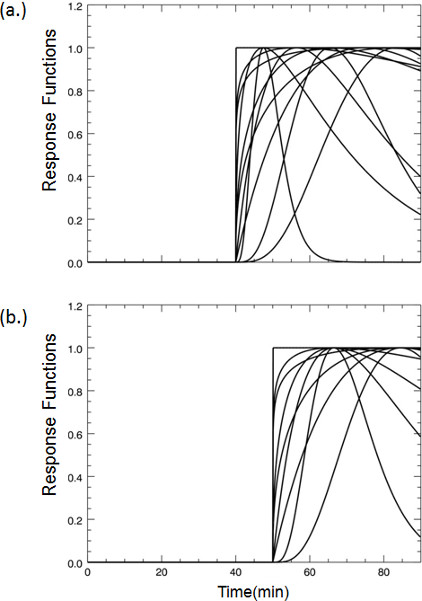
Figure 3. Examples of dopamine response functions that take-off at (a) 40 min or (b) 45 min post-tracer injection.
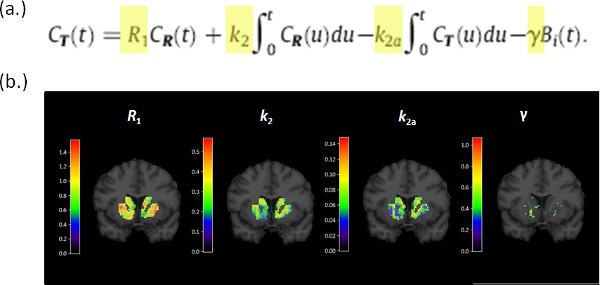
Figure 4. Parametric images generated by fitting the lp-ntPET operational equation (a) to the PET data. (b) Images corresponding to the 4 parameters of the model, R1, k2, k2a, γ, are evaluated for the striatum and shown overlaid on the corresponding MR slice. Click here to view larger figure.
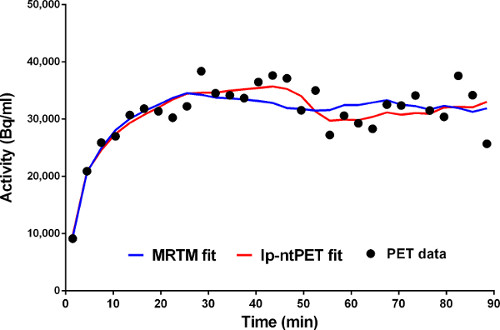
Figure 5. Fits of MRTM (blue) and lp-ntPET (red) models to time-activity data from a single voxel.
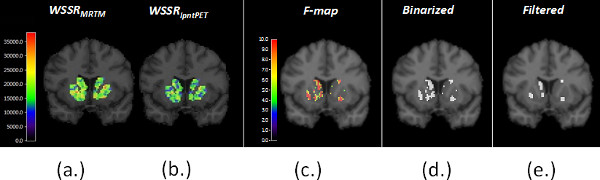
Figure 6. Parametric images of the WSSR for (a) MRTM and (b) lp-ntPET. The respective WSSR maps are compared to create the F-map (c), which in turn is thresholded to a binary mask (d) and then filtered to produce the Final Significance Mask. Click here to view larger figure.
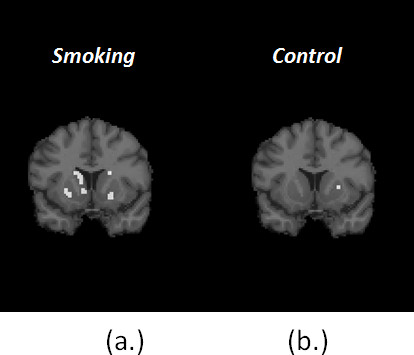
Figure 7. Comparison of Final Signifcance Masks for smoking (a) and control (b) conditions for the same subject.
Figure 8. Single-slice dopamine movie for single subject in control (‘Rest’) and smoking conditions. Click here to view Figure 8.
Figure 9. Multi-slice dopamine movie for single subject in (top) smoking and (bottom) control (‘Rest’) conditions. Click here to view Figure 9.
